Memoria de trabajo y consciencia: tres perspectivas teóricas1
Working Memory and Consciousness: Three Theorical Frameworks
Memória de trabalho e consci ência: tr ês quadros teóricos
Recibido el 26 de mayo de 2021. Aceptado el 26 de febrero de 2022
› Para citar este artículo: Landínez Martínez, D., Montoya Arenas, D. A. y Pineda, D. A. (2022). Memoria de trabajo y consciencia: tres perspectivas teóricas. Ánfora, 29(53), 244-272. https://doi.org/10.30854/anf.v29.n53.2022.800
La perspectiva de la Memoria de Trabajo (MT) como proceso consciente ha permitido definir la consciencia como el contenido de la MT; sin embargo, han surgido inquietudes sobre las comparaciones que se han realizado entre ambas. Objetivo: aunque la relación entre estos dos campos de estudio ha sido planteada desde la psicología, la filosofía y la neurociencia, una revisión teórica que aborde los elementos centrales de las perspectivas más citadas enriquecería el debate en esta área de conocimiento. Metodología: esta revisión se centra en tres perspectivas teóricas: 1) el modelo multicomponente de memoria de trabajo; 2) la Teoría del Espacio de Trabajo Global (GWT); 3) el modelo jerárquico. Se analizaron 113 artículos en los que se abordaron las tres perspectivas anteriores. Resultados: el modelo multicomponente de memoria de trabajo aporta una descripción funcional básica sobre cómo las representaciones mentales permanecen en línea durante un procesamiento cognitivo complejo. De este modo, de un lado, el intercambio de información entre el ejecutivo central y el búfer episódico; y de otro lado, el bucle fonológico y la agenda visuoespacial se presenta a través del procesamiento consciente. Conclusiones: asimismo, el ejecutivo central controla y modifica la atención, pero el búfer episódico permite la disponibilidad de información multimodal.
The view of Working Memory (WM) as a conscious process has allowed defining consciousness as the content of working memory. However, concerns have emerged over comparisons between consciousness and working memory. Goal: although the relationship between these two study fields has been the matter of psychology, philosophy and neuroscience, a theoretical review addressing the core elements of highly cited perspectives would enrich the discussion in this study area. Method: this review focuses on three theoretical frameworks: 1) the multi-component model of working memory, 2) the global workspace theory, 3) the hierarchical framework. The authors analyzed 113 articles which discussed the previous three models. Results: the multi-component model of working memory contributes a basic functional description on how mental representations remain on-line during complex cognitive processing. Thereby, the information exchange between the central executive and the episodic buffer, in one sense, and the phonological loop and the visuo-spatial sketchpad in the other is given through conscious processing. Conclusions: likewise, the central executive controls and changes attention but the episodic buffer allows multimodal information availability.
A visão da Memória de Trabalho (WM) como um processo consciente permitiu definir a consciência como o conteúdo da memória de trabalho. No entanto, surgiram preocupações sobre as comparações entre consciência e memória de trabalho. Objetivo: embora a relação entre estes dois campos de estudo tenha sido a questão da psicologia, filosofia e neurociência, uma revisão teórica abordando os elementos centrais de perspectivas altamente citadas enriqueceria a discussão nesta área de estudo. Metodologia: esta revisão se concentra em três estruturas teóricas: 1) o modelo multicomponente de memória de trabalho, 2) a teoria do espaço de trabalho global, 3) a estrutura hierárquica. Os autores analisaram 113 artigos que discutiam os três modelos anteriores. Resultados: o modelo multicomponente de memória de trabalho contribui com uma descrição funcional básica sobre como as representações mentais permanecem on-line durante o complexo processamento cognitivo. Assim, a troca de informações entre o executivo central e o buffer episódico, em um sentido, e a alça fonológica e o bloco de desenho visuoespacial no outro se dá por meio de processamento consciente. Conclusões: da mesma forma, o executivo central controla e muda a atenção, mas o buffer episódico permite a disponibilidade de informações multimodais.
Según la teoría de la información integrada (Tononi y Koch, 2015), la consciencia es la capacidad del cerebro para integrar la información de manera rápida; es un sintetizador de contexto holístico en el que el individuo está inmerso. Este tipo de capacidad requiere de un sistema tálamocortical que genere oscilaciones en frecuencia gamma (Tononi et al., 2016). De hecho, las lesiones tálamocorticales están altamente relacionadas con la pérdida global de la consciencia, tal y como se observa después de un coma (Laureys et al., 2004). La actividad neuronal correlacionada con la experiencia consciente está ampliamente distribuida sobre la corteza, lo que indica que la consciencia depende de la red tálamocortical en lugar de un área cortical única (Tononi et al., 2016). Esta afirmación sugiere que las lesiones en regiones corticales específicas pueden afectar la experiencia consciente sin afectar la consciencia global, como la incapacidad de percibir rostros (Kolb y Whishaw, 2006).
El Correlato Neuronal de la Consciencia (NCC por sus siglas en inglés) se define como el conjunto mínimo de mecanismos neuronales suficientes para cualquier percepción consciente específica (Crick y Koch, 1990; Frith, 2005). Esta definición supone dos posibles interpretaciones, según el contenido específico de la consciencia o el estado general de ser consciente.
El contenido específico de la consciencia son los mecanismos neuronales que determinan una característica particular dentro de la experiencia. Por ejemplo, el NCC para experimentar el contenido específico de un rostro, corresponde a las neuronas que se activan, cada vez que un individuo observa, imagina o sueña un rostro, y que permanecen inactivas bajo otras circunstancias (Frith, 2005). Cuando estas neuronas se activan mediante Estimulación Magnética Transcraneal (EMT), un individuo debería ver un rostro, aunque no aparezca ninguno en pantalla; por el contrario, si la actividad de dichas neuronas se bloquea, no debería ver ningún rostro, aunque aparezca en pantalla (Koch et al., 2016). Desde una perspectiva diferente, el NCC es el sustrato neuronal que estimula la experiencia consciente como un todo (Koch et al., 2016).
La investigación en torno al NCC ha permitido la identificación de descargas neuronales sincronizadas sobre la corteza visual como respuesta a dos estímulos visuales que generan ondas gamma (30-70 Hz); sin embargo, esto ha suscitado preguntas y dudas en la comunidad científica (Gray et al.,1989). Hoy en día se acepta que la consciencia requiere que las neuronas estén sincronizadas a través de descargas rítmicas (ondas gamma) (Crick y Koch, 1990) para explicar la integración de múltiples estímulos en una sola experiencia (Singer, 1999). La sincronización sobre la corteza visual se da por medio de la atención (Roelfsema et al., 1997) y la estimulación del sistema reticular activador (Herculano-Houzel et al., 1999; Roelfsema et al., 1997); asimismo, refleja la dominancia perceptiva en tareas de rivalidad binocular, aunque las tasas de activación no cambien (Fries et al., 1997). Estudios previos con electroencefalografía y magnetoencefalografía sugieren que la sincronía gamma también se correlaciona con la consciencia visual (Melloni et al., 2007; Rodriguez et al., 1999).
No obstante, la mayoría de los estudios no diferenciaron la visibilidad consciente de la atención selectiva. Una vez que esto ha ocurrido, la sincronización de las ondas gamma de rango amplio se correlaciona con la atención, tanto si el estímulo fue visto como si no, pero las ondas gamma de rango medio se correlacionan con la visibilidad del estímulo (Wyart y Tallon-Baudry, 2008). La sincronía gamma podría aumentar durante el sueño NMOR en pacientes anestesiados (Imas et al., 2005; Murphy et al., 2011) o con convulsiones (Pockett y Holmes, 2009) o incluso cuando se exponen a estímulos que provocan respuestas emocionales inconscientes (Luo et al., 2009). Estos resultados indican que la sincronía gamma podría presentarse cuando hay falta de consciencia. Un estudio anterior utilizó electrocorticografía sobre la corteza visual y demostró que las oscilaciones gamma de baja amplitud se generan mediante patrones espaciales (rejillas luminosas), pero no con ruido o imágenes que puedan verse (Hermes et al., 2015). Por último, estos resultados sugieren que las ondas gamma no son totalmente necesarias a la vista (Ray y Maunsell, 2011).
Otros marcadores electrofisiológicos para explorar la consciencia son los Potenciales Relacionados con Eventos (PREs), específicamente la respuesta que ocurre alrededor de los 300 ms (P3b) después de la presentación del estímulo. El componente P3b es provocado por estímulos visuales o auditivos y se presenta sobre la región frontoparietal (Sutton et al., 1965). Algunos estudios con tareas basadas en paradigmas han demostrado que el P3b es un correlato neuronal para el reporte de estímulos (Dehaene y Naccache, 2001; Del Cul et al., 2007; Sergent et al., 2005). Por lo tanto, el componente P3b que se mide a través del paradigma oddball auditivo se ha propuesto como una señal precisa de la consciencia sobre la red frontoparietal (Dehaene y Changeux, 2011).
Pese a ello, esta perspectiva se ha subestimado a lo largo de los estudios experimentales. Por ejemplo, un estímulo irrelevante para la ejecución de una tarea, no activa el componente P3b (Silverstein et al., 2015); incluso si el individuo es completamente consciente de ello (Pitts et al., 2014). No obstante, aquellos estímulos de los que el individuo no es consciente pueden activar dicho componente (Silverstein et al., 2015). Lo anterior no corresponde a una señal de percepción consciente, una vez que el estímulo ya está en la memoria de trabajo (MT). (Melloni et al., 2011). Por otro lado, un PRE que comienza a los 100 ms después de la presentación del estímulo alcanzará su pico alrededor de los 200-250 ms. Este se localiza en la corteza posterior (Pitts et al., 2014; Railo et al., 2011) y se correlaciona con la percepción consciente.
Las descargas rápidas de bajo voltaje en el Electroencefalógrafo (EEG) mientras se está despierto, también conocidas como EEG activo (Moruzzi y Magoun, 1949), fue uno de los primeros índices de consciencia y continúa siendo uno de los mejores marcadores sensibles. Los registros intracelulares de las neuronas corticales y talámicas de los gatos han develado los mecanismos subyacentes de la transición de las descargas rápidas de bajo voltaje (despierto) a las descargas lentas de alto voltaje comunes durante el sueño profundo o incluso bajo anestesia (Steriade, 2000). Cuando las neuronas talámicas se hiperpolarizan, pasan de un modo tónico a un modo de disparo rápido que da lugar a la sincronización del EEG en ondas theta (5-7 Hz) (Steriade, 2000). Las oscilaciones más amplias de las ondas delta se observan cuando las neuronas corticales comienzan a alternar entre estados de despolarización e hiperpolarización cada segundo (Steriade et al., 2001).
Este grupo de cambios fisiológicos se presenta junto con la pérdida de consciencia bajo condiciones farmacológicas, patológicas y fisiológicas (Brown et al., 2010). Entre las estrategias más precisas para evaluar la pérdida de consciencia, la mejor opción es la detección de ondas lentas de gran amplitud (Kertai et al., 2012; Murphy et al., 2011). Por ejemplo, las ondas lentas son las más comunes durante el sueño profundo en las primeras horas de la noche; una vez que los individuos son despertados, niegan cualquier percepción (Siclari et al., 2013). De igual manera, un aumento repentino de la potencia de la onda coincide con la pérdida de consciencia cuando se induce la anestesia por medio de Propofol (Murphy et al., 2011; Purdon et al., 2013). En un entorno clínico, un cambio de patrón de ondas delta a alfa explica la transición del estado vegetativo al estado de mínima consciencia (Schiff et al., 2014).
Ante estos hallazgos previos, algunos estudios plantean que la consciencia es una forma de procesar información y establecen la importancia del NCC (Aleksander, Gamez, 2011; Baars, 1988; Earl, 2014; Fingelkurts y Fingelkurts, 2017; Tononi, 2012). La consciencia permite la transmisión y el procesamiento de la información. Ahora bien, como la información que produce es significativa para el individuo que la experimenta conscientemente, este sabe lo que significa. Por último, la información que produce es «individualizada» (Jonkisz, 2016) en tanto tiene «ese» significado solo para quien la experimenta, y no para otros individuos. Por ejemplo: Yo sé lo que significa para mí experimentar «miedo», pero otro individuo no puede saber de manera directa lo que esto significa para mí (y viceversa).
Por lo tanto, una cuestión relevante cuando se analiza la consciencia, es abordar el yo, entendido como un mecanismo que permite la individualidad y la continuidad de la experiencia consciente. Así es como el yo adquiere importancia para el organismo, conforma el cuerpo y la mente en una sola unidad: la personalidad. Así, el yo es una característica autobiográfica, que se actualiza de manera constante y, al mismo tiempo, se expande sobre las representaciones subjetivas de la realidad, incluso sobre el escenario emocional. Este dispositivo posee un correlato fisiológico que se entiende en la representación del cuerpo (circuitos neuronales), sus proyecciones al hipocampo, la amígdala y la corteza prefrontal medial (Arturo et al., 2015; Damasio, 2003a; Damasio, 2003b). Para que esto ocurra, el yo consciente siempre necesita almacenar y actualizar cada experiencia en la memoria. Esta perspectiva considera a la MT como el dominio cognitivo básico que subyace a la consciencia (Marchetti, 2018).
La MT es un sistema cognitivo que participa en el almacenamiento y procesamiento temporal de la información para poder realizar una tarea (Baddeley, 2017). Asimismo, cuando se realiza una tarea cognitiva, la MT discrimina entre información relevante y no relevante (Unsworth y Engle, 2007); también participa activamente en el control de la atención (Broadway y Engle, 2011). Este punto de vista ha permitido considerar la MT como un «espacio de trabajo» donde ocurre el pensamiento y la cognición (Baars y Franklin, 2003). Tal afirmación se ha comprobado empíricamente dada la asociación consistente entre la MT y los procesos cognitivos superiores (Engle, 2002).
La relación entre MT y consciencia es evidente en el modelo multicomponente de Baddeley, el cual es relevante para el presente estudio. (Baddeley, 2017). Por ejemplo, se supone que la información que permanece en la memoria de trabajo es consciente. Según lo anterior, la MT opera con información a la que se accede conscientemente y, de alguna manera, se asume que la MT y la consciencia son lo mismo. Esta perspectiva, que es discutible, puede justificar una mejor definición de lo que es la consciencia más allá del mero contenido de la MT; sin embargo, han surgido dudas respecto a la idea de que la MT sea comparada con la consciencia, dado que ambos conceptos son unidades integradas (Cowan, 2012).
Aunque la consciencia y la MT comparten propiedades funcionales, la forma en la que se relacionan entre sí es de gran interés. Por ejemplo, un campo de investigación se centra en cómo el contenido de la MT puede afectar a la consciencia. Esta perspectiva aborda cómo las representaciones visuoespaciales se mantienen en línea y evalúa la consciencia a través de la supresión de estímulos (Jiang et al., 2007). Al realizar esta tarea, un grupo de estímulos proyectados hacia uno de los ojos suprime la visibilidad de un estímulo fijo que se presenta simultáneamente al otro ojo (Tsuchiya y Koch, 2005). Por último, cada individuo debe indicar una vez que sea consciente del estímulo fijado, lo que suele tardar un par de segundos.
Un ejemplo claro de lo anterior es un estudio en el que los pacientes debían recordar un color mientras realizaban una tarea de detección de círculos (Gayet et al., 2013). Estos hallazgos demostraron que el contenido de la MT visuoespacial activa un umbral de alerta. Este umbral fue más bajo cuando el color de los círculos era similar al que se mantenía en línea en la MT. También se encontró que el umbral de consciencia fue menor cuando el rostro presentado en pantalla era similar al rostro mantenido en línea en la MT (Pan et al., 2014). En conjunto, estos estudios sugieren que el contenido de la MT visuoespacial puede influir en el umbral de consciencia visual, pero también sesga el acceso a la información consciente.
Otra línea de investigación ha relacionado la consciencia con varios procesos de la MT y con la actividad prefrontal. Por ejemplo, en un estudio se les solicitó a los participantes aprender un grupo de letras mientras realizaban una tarea de enmascaramiento en la que tenían que detectar un dígito específico (De Loof et al., 2013). Los resultados mostraron que la tarea de detección tuvo puntuaciones más bajas cuando el número de letras había aumentado. Esto indicaría un aumento del umbral de consciencia una vez que la información llega a la MT (Lavie, 2005).
Los hallazgos anteriores indican que la MT incide en la consciencia; no obstante, la relación entre ambas aún no está clara. En la siguiente sección se explicarán tres marcos teóricos que abordan esta relación. Aunque la MT se explica desde diversas perspectivas, este artículo de revisión no abordará el modelo de procesos embebidos de Cowan (2012), ya que su enfoque principal es enfatizar los vínculos entre la memoria y la atención. Desde este punto de vista, los estímulos con características físicas que han permanecido relativamente inalteradas a lo largo del tiempo y que no tienen una importancia relevante para el individuo, siguen activando características en la memoria, pero no suscitan consciencia.
Otro enfoque para explicar la MT proviene de Oberauer, quien definió la MT como un medio para construir, mantener y manipular representaciones temporales que controlan los pensamientos y acciones en curso (2009); sin embargo, al igual que el modelo de Cowan, este enfoque mantiene el papel de la atención como mecanismo de selección. En este sentido, la MT es una forma de atención: los contenidos de la MT se seleccionan en tanto son relevantes para la tarea en curso. A menudo, las diferentes teorías—de la MT o de otro tipo— no pueden compararse directamente porque las teorías, aunque nominalmente tratan el mismo tema, en realidad se basan en definiciones sutilmente diferentes de lo que se estudia (Alloway et al ., 2005) .
Dado que este artículo pretende revisar la estrecha relación entre MT y consciencia, se presentarán otros enfoques que han introducido explícitamente este interés. En la última sección, se propone una línea de investigación para futuros estudios.
El modelo de MT de Baddeley sugiere una organización jerárquica compuesta por el ejecutivo central y algunos sistemas esclavos para el almacenamiento de la información (Baddeley, 2017). Los sistemas esclavos permiten almacenar temporalmente información específica de cada modalidad: verbal (bucle fonológico), visual, espacial (agenda visuoespacial). Asimismo, el acceso a los sistemas esclavos se considera consciente. De hecho, un individuo es consciente del repaso articulatorio de la información almacenada en el bucle fonológico.
Un hecho arbitrario sobre el bucle fonológico es que el propio repaso articulatorio no tiene capacidad de almacenamiento interno. En otras palabras: «la voz interior no se escucha a sí misma» o «la voz interior es sorda» (Buchsbaum, 2013). Por otro lado, si se piensa en el repaso articulatorio como un mecanismo que almacena y reactiva su propio contenido, este se vuelve autosuficiente desde la teoría del procesamiento de la información: es una voz interior que se puede escuchar.
Dejando de lado estas consideraciones conductuales, a favor o en contra de la arquitectura del bucle fonológico, parecería que estas estuvieran en contra de la evidencia científica que afirma: el habla interior es una versión privada del habla exterior. Por lo tanto, las cualidades auditivas del oído interno son bastante similares a la escucha del habla externa. Por ejemplo, desde una perspectiva fenomenológica, una vez que se ha imaginado un punto verde, es como observar el mismo estímulo (Shepard y Chipman, 1970; Smart, 1995). Del mismo modo, durante el habla interna, la información verbal está relacionada con el contenido de una imagen auditiva en el oído interno, por lo que puede ser reportada conscientemente.
Sin embargo, no es el mismo caso para la voz interior: aunque alguien puede reportar un sentimiento de agencia durante el habla interior (Morsella et al., 2011), este sentimiento no tiene contenido lingüístico y no hay otros tipos de sentimientos que puedan ser descritos como la representación de un mensaje verbal. En consecuencia, un análisis introspectivo del discurso interior favorece la existencia de dos componentes independientes y conscientes.
La voz interior es un conocido marcador de agencia que expresa la idea: «Eres tú quien habla», pero el oído interno lleva el contenido consciente del mensaje: «Esto es lo que dices». De hecho, la experiencia consciente de la conducta interior carece de contenido, salvo algunos marcadores de agencia como los impulsos, los planes y las intenciones (Morsella et al., 2011). Para poder acceder conscientemente al programa motor del habla, la representación de la conducta debe ser capaz de proyectarse al espacio sensorial-perceptivo. Por lo tanto, podría afirmarse que el contenido del programa motor no es vulnerable a la introspección sin ser completado. Esta es una cualidad necesaria de un organismo autoconsciente: ser capaz de anticipar una conducta una vez que la haya ejecutado o al menos simulado internamente (Libet et al., 1982). Una forma más de entender el acceso al contenido del programa motor es asumir que las representaciones conscientes son independientes entre sí. En otras palabras, una representación no puede observarse a sí misma.
De otro lado, el estado consciente del ejecutivo central está menos claro. Este se centra en el procesamiento cognitivo controlado (Atkinson y Shiffrin, 2016), por lo que debe ser consciente. Los procesos ejecutivos conscientes parecen estar relacionados con el cambio voluntario de la atención o la inhibición de la distracción de los estímulos irrelevantes para realizar una tarea. Este modelo de MT también relaciona la experiencia consciente con el ejecutivo central (Baddeley, 1992). La evidencia empírica se observa cuando la supresión de la ensoñación no depende de una sola modalidad; depende únicamente del ejecutivo central. Dado que la ensoñación es un estado en el que la mente es completamente independiente, el ejecutivo central se ha convertido en el componente de la MT que accede a la información consciente (Baddeley, 1992).
Pese a ello, los procesos ejecutivos, como las estrategias de búsqueda en la memoria a largo plazo, no están bajo control consciente. En consecuencia, el acceso consciente al ejecutivo central es parcial (Velichkovsky, 2017).
Como cada sistema esclavo de la MT se consideraba específico según la modalidad, posteriormente, el modelo se actualizó con la inclusión del búfer episódico para mantener en línea los episodios multimodales (Baddeley, 2000). Los episodios del búfer son el resultado de la integración sensorial, los recuerdos y la imaginación. Por un lado, el búfer episódico interactúa con los sistemas de almacenamiento de modalidad única y, por otro, con la memoria episódica a largo plazo. Así es como el búfer episódico vincula la memoria y la consciencia (Baddeley, 2000); además, es un almacén pasivo que contiene unidades de información integradas (chunks) y que permite al ejecutivo central acceder a la consciencia (Baddeley et al., 2010).
En resumen, el sistema de almacenamiento alberga representaciones experimentadas conscientemente. El búfer episódico contiene unidades integradas multimodales para poder abarcar la experiencia consciente; sin embargo, es el ejecutivo central el que interactúa con la consciencia y permite el acceso consciente a aquellos estímulos que permanecen en la MT (Baddeley, 2010).
El modelo multicomponente de MT ha introducido una de las teorías más sólidas para respaldar una MT consciente. De hecho, se supone que todas las memorias de trabajo operan con un contenido consciente. El ejecutivo central es un sistema consciente debido a que lleva a cabo el control consciente sobre la MT y la atención. Los propios sistemas de almacenamiento representan conscientemente su contenido, como es evidente en el bucle fonológico y en la agenda visuoespacial. Una vez que los estímulos se han disipado de la consciencia, se supone que han salido completamente de la MT (Velichkovsky, 2017).
Según lo anterior, no es una tarea fácil conciliar la idea de una MT inconsciente con el modelo multicomponente. Parece ser el reto dentro del modelo de Baddeley dado que, por un lado, se propone una diferencia entre la MT y la memoria a largo plazo y, por otro, se postula la diferencia entre el procesamiento cognitivo controlado y el automático. Entender la MT inconsciente en el modelo multicomponente es una línea de investigación futura. Se requerirán cambios en el modelo actual, tales como la incorporación de los estados de representación al sistema esclavo (Velichkovsky, 2017).
Un modelo ampliamente aceptado que aborda la estrecha relación entre la MT y la consciencia es la Teoría del Espacio de Trabajo Global (GWT por sus siglas en inglés) (Baars, 2005). Desde esta perspectiva, (1) el cerebro puede configurarse como un sistema masivo de procesamiento paralelo, de modo que (2) los procesadores cerebrales pueden trabajar juntos mediante el intercambio de datos. El papel de la consciencia es la disponibilidad de datos para los procesadores cerebrales. Un ejemplo común de datos compartidos a través del espacio de trabajo global es la entrada sensorial. La GWT se basa en la evidencia científica que afirma que la experiencia consciente implica múltiples activaciones a lo largo del cerebro (red frontoparietal) en comparación con estados inconscientes como el sueño y el coma (actividad interhemisférica disminuida) (Baars, 2005).
La GWT también se conoce como la metáfora de la consciencia donde los actores (datos) en un escenario (MT) son iluminados por un foco (atención) (Baars y Franklin, 2003). La zona oscura del escenario está relacionada con aquellos aspectos de la memoria inmediata que no son el centro de atención. El trabajo realizado por los actores es controlado «entre bastidores» por procesos ejecutivos como el yo (Baddeley, 2017). Finalmente, toda la escena es observada por una audiencia inconsciente compuesta por sistemas motivacionales y procesos automáticos (Baddeley, 2007).
Tanto la GWT como el modelo de MT parecen similares, de hecho, se ha afirmado que varios procesos de la MT cumplen con los principios de la GWT, lo que establece una estrecha relación entre ambas teorías (Velichkovsky, 2017). Pese a ello, un análisis más exigente de la GWT revela las diferencias entre la MT y la consciencia. Desde esta perspectiva, la MT asume varios procesos inconscientes (Bergström y Eriksson, 2014). Por ejemplo, se supone que el almacenamiento preconsciente de información sobre los sistemas esclavos es específico de la modalidad. Entonces, la interacción cognitiva entre la MT y la consciencia podría explicarse de la siguiente manera: una vez que la entrada sensorial ha sido registrada, es enviada a través de los procesos atencionales a los procesadores cerebrales para convertirse en consciente. El contenido de la consciencia se mantiene en el bucle fonológico (Soto et al., 2011). Por consiguiente, los planes de acción y los objetivos empiezan a funcionar una vez que se opera sobre datos conscientes. Asimismo, estos planes están bajo la supervisión de la MT que opera de forma inconsciente. Por ello, las operaciones conscientes abarcan una sola parte del ciclo cognitivo (Velichkovsky, 2017).
Parece que la consciencia como foco atencional es esencial para entender mejor la relación entre la consciencia y la MT en la GWT (Baars, 2005). Este foco elige una fracción de las representaciones cognitivas a las que accederán los procesadores cognitivos inconscientes. La disponibilidad universal de datos permite a los procesadores cerebrales empezar a planificar una tarea bajo poco control consciente (Hassin et al., 2009). En consecuencia, el objetivo de la consciencia es recoger las representaciones internas dentro del contenido de la MT. Esta idea convierte la consciencia en un proceso de atención interior. De esta manera, la relación entre la consciencia y la MT, desde esta perspectiva, es inclusiva. La consciencia se incorpora a la MT y está compuesta por procesos motores, ejecutivos y perceptivos inconscientes. Por lo tanto, la consciencia es un subconjunto de la MT (Baars, 1988).
Desde su desarrollo, se han planteado algunas inquietudes en torno a la GWT respecto a la descripción del papel de la consciencia en las conductas dirigidas a objetivos (Baars y Franklin, 2003). Esta teoría no presenta los mecanismos internos de la MT, lo que dificulta la explicación de cómo el contenido consciente e inconsciente del modelo interactúa. Lo anterior sugiere una futura línea de investigación relacionada con las diferencias cualitativas entre dicho contenido consciente e inconsciente de la MT (Velichkovsky, 2017).
Se publicaron una serie de artículos sobre el estudio de los lóbulos frontales (Burgess y Stuss, 2017). Como resultado, se pueden identificar tres escenarios de estudio sobre la función de dichos lóbulos: (1) desarrollo de pruebas para evaluar los síntomas ejecutivos, (2) conocimientos actuales en neuroanatomía, (3) estudios que utilizan técnicas de neuroimagen para comprender de mejor manera el cerebro humano (Burgess y Stuss, 2017). Por último, se propusieron tres niveles de organización jerárquica del cerebro.
El primer nivel está relacionado con la actividad cerebral compartida de los sistemas funcionales (Stuss y Alexander, 2005). Su correlato neuroanatómico está sobre la corteza frontal medial y dorsolateral. Estas dos áreas cerebrales parecen permitir: (a) la organización de la información en una secuencia significativa y (b) las habilidades para dirigir el comportamiento. El segundo nivel de funcionamiento del lóbulo frontal está relacionado con el control; su base anatómica es la corteza prefrontal ( Andrs y Van der Linden, 2001). Esta función de control está relacionada con focalización consciente en un objetivo y puede dividirse en varias funciones psicológicas: anticipación, selección de objetivos, planificación y monitoreo (Crick y Koch, 2003). El último nivel de la función frontal es la consciencia y la autoconsciencia. El correlato neuroanatómico está sobre la corteza prefrontal que también está cerca de la metacognición (Stuss y Alexander, 2005).
Sin embargo, un estudio posterior sobre este modelo permitió establecer tres procesos cognitivos relacionados con el desempeño en tareas ejecutivas: energización, vista como el proceso de iniciar y mantener una respuesta; configuración de la tarea, que se refiere a la capacidad de establecer una relación estímulo-respuesta para responder a un objetivo con atributos específicos (planificación y organización); monitoreo, entendido como el proceso de comprobación de una tarea a lo largo del tiempo; y para el control de calidad (Burgess y Stuss, 2017). Lo más probable es que el monitoreo sea el proceso cognitivo más cercano a la MT porque puede asumirse como una supervisión en línea. También es una de las habilidades más importantes relacionadas con la regulación metacognitiva (Schraw y Dennison, 1994).
Finalmente, el monitoreo metacognitivo implica un proceso de calibración en el que un sujeto monitorea su propio proceso de pensamiento y estado de conocimiento a través de cada sistema esclavo de la MT (Schraw y Moshman, 1995).
A continuación, se presentan algunos ejemplos clínicos sobre las etapas anteriores:
Energización: un individuo con daños en la corteza dorsomedial presenta una serie de déficits específicos, tales como velocidad de procesamiento lenta. Por ejemplo, este individuo no podría mantener la información en línea (lista de palabras) durante los últimos 45 segundos de una tarea de fluidez verbal en comparación con los primeros 15 segundos. Esto se debe a la incapacidad de iniciar y mantener una respuesta (Picton et al., 2007).
Configuración de la tarea: un individuo con daño en el hemisferio izquierdo aumentará la tasa de falsos positivos en tareas como el test de Stroop. Este déficit es bastante común durante las primeras etapas del aprendizaje (Stuss y Knight, 2009).
Monitoreo: un individuo con daño en el hemisferio derecho aumentará la tasa de falsos positivos y se le dificultará realizar tareas aritméticas (cálculo, recuperación, uso de estrategias, toma de decisiones) (Stuss, 2011).
Probablemente, la anosognosia es uno de los casos clínicos más conocidos en los que un individuo ve disminuidos sus niveles de consciencia. Se trata de un déficit de dominio específico y está relacionado con un único sistema funcional en el que la falta de consciencia parece ser la ausencia de conocimientos factuales (Stuss, 1991).
Los pacientes con una lesión cerebral focal en la región posterior del hemisferio derecho muestran varios déficits como: hemianopsia izquierda, prosopagnosia y hemi-inatención (Stuss, 2011). Si esta lesión se extiende a las zonas anteriores, el paciente presentaría una hemiplejía izquierda. Asimismo, la anosognosia también puede interpretarse como un déficit secundario relacionado con la conservación de las capacidades necesarias para realizar las actividades básicas de la vida diaria.
El yo es uno de los sistemas cognitivos básicos que subyacen a la experiencia consciente. Se desarrolla sobre valores biológicos, naturalmente seleccionados y culturalmente adquiridos. Es el motor que permite sostener y ampliar el bienestar de un individuo en su conjunto. El yo se expresa a través del sistema nervioso central y periférico, que mapea el cuerpo, su contexto y el entorno. Es a través del yo que la compleja estructura de un organismo se convierte en la voz de un solo individuo (Marchetti, 2018).
Un amplio conjunto de valores rige el yo. Un individuo puede sentirse abrumado por múltiples estímulos internos y externos. El cuerpo se ajusta constantemente al entorno. Esto requiere un mecanismo que, por un lado, permita enfocarse en los datos más importantes para lograr un objetivo según el contexto, pero, por otro lado, inhibe el ruido de los estímulos irrelevantes. Este mecanismo se conoce como atención (Marchetti, 2018).
La atención permite a un individuo controlar grandes cantidades de información, pero también le permite seleccionar los datos más importantes para cumplir un objetivo (Awh et al., 2012). Esto puede lograrse de múltiples maneras: a través de un procesamiento ascendente exógeno y no intencionado; a través de un mecanismo descendente endógeno y voluntario (Carrasco, 2011; Chica et al., 2013); dirigiendo interna o externamente el foco de atención (Corbetta et al., 2008; Corbetta y Shulman, 2002); y manteniendo la atención durante un tiempo limitado (Zeman, 1996), incluso cuando se ha distribuido en múltiples estímulos (Eimer y Grubert, 2014).
La atención es una herramienta precisa para la resolución de problemas, de hecho, permite dividir el flujo de información en episodios atencionales y cada episodio solo considerará el contenido de un subconjunto momentáneo de problemas (Duncan, 2013). Por último, el proceso de selección atencional establece nuevas dimensiones experimentales además de las establecidas anteriormente. Al seleccionar estímulos no relacionados, se hace posible la simulación de nuevos escenarios que un individuo podría no haber experimentado conscientemente si no dispusiera de capacidades atencionales (Marchetti, 2018). Por lo tanto, la consciencia humana es capaz de utilizar el poder del cerebro para construir el pensamiento secuencial y hacer simulaciones mientras está despierto sin necesidad de estímulos sensoriales (Baumeister y Masicampo, 2010).
Aunque el papel de la atención puede asumirse teóricamente como un proceso ininterrumpido y continuo que permite cambiar de tarea, los datos actuales apoyan la idea de que la atención actúa periódicamente. Esta periodicidad es producto de las oscilaciones cerebrales (Marchetti, 2018). La atención proporciona una plantilla para dar forma a las experiencias conscientes, por consiguiente, estas experiencias se ajustan al papel de la atención para detectar cambios en el yo.
No obstante, la atención por sí misma no es suficiente para que se produzca la experiencia consciente más compleja. La atención se encarga de seleccionar los elementos centrales de la experiencia consciente, pero sigue siendo necesario un mecanismo como la MT para combinar y ensamblar estos elementos (Marchetti, 2018).
La MT no solo consiste en recordar información, sino que es una habilidad más general relacionada con el control atencional que ejerce un control descendente sobre la cognición. La MT también permite combinar secuencialmente los estímulos. Una de las principales funciones de la MT es mantener las representaciones a través de la unión temporal entre el contenido (estímulos, palabras) y el contexto (localización del estímulo en una tarea visuoespacial) (Oberauer, 2009). Del mismo modo, la unión también se ha relacionado con las características declarativas y procedimentales de la MT.
Algunos trabajos que utilizan técnicas neurofisiológicas como el EEG han empezado a centrarse en este sistema. Los estudios han encontrado que para que el cerebro sea capaz de codificar, retener y recuperar información a través de la MT, necesita sincronizar varias tareas con el apoyo de conjuntos neuronales locales que trabajan en diferentes escalas temporales anidadas en la misma jerarquía operativa (Fingelkurts et al., 2010; Monto, 2012). Especialmente, los módulos operativos (span) parecen ser necesarios para una memoria exitosa (Fingelkurts et al., 2003). De hecho, aunque la codificación y la recuperación de la memoria comparten regiones comunes en la corteza cerebral, la sincronía operativa de cada tarea se ve como un grupo de módulos anidados a las tareas de memoria a corto plazo (Fingelkurts et al., 2003). Tanto un número grande como uno pequeño de módulos operativos podría causar un déficit de memoria.
De este modo, la interacción entre las ondas de la banda theta (4-8 Hz) y las ondas de la banda gamma (30-200 Hz) es la característica principal de los estímulos ordenados secuencialmente en la MT (Lisman y Jensen, 2013). Una vez integrada la información, la MT permite combinar y ensamblar la información en la consciencia (Marchetti, 2014).
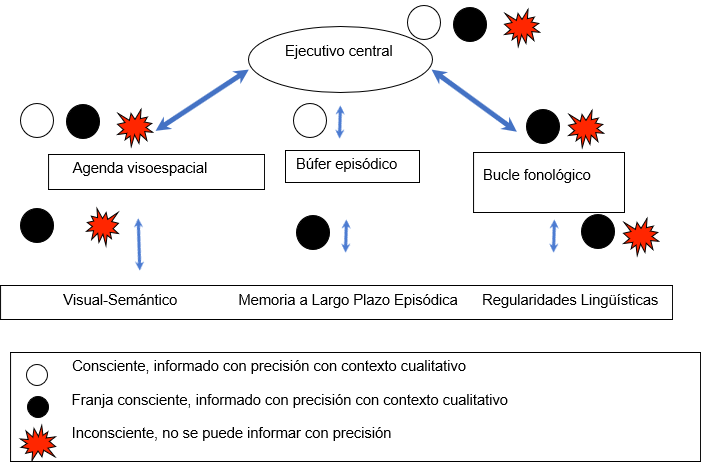
El modelo de MT de Baddeley es quizás uno de los más influyentes en la cognición de las últimas décadas (Andrade, 2002; Baddeley, 2017). Cada uno de los componentes es fácilmente evaluable, como el bucle fonológico (repetición de palabras o números), la agenda visoespacial (imágenes mentales) y el ejecutivo central (manipulación voluntaria). La figura 1 muestra que la MT está fuertemente involucrada con la consciencia, tanto desde una perspectiva cualitativa (discurso interno), o experiencias conscientes como la intención de repasar estímulos previamente almacenados en la MT (Baars y Franklin, 2003). El yo participa tanto en el ejecutivo central como en el bucle fonológico y permite la individualidad y la continuidad de la experiencia consciente. Así es como el yo se convierte en una referencia para un organismo compuesto de cuerpo y mente. Una unidad indivisible: Personalidad.
Figura 1. Presentación esquemática del Modelo de Memoria de Trabajo. Fuente: (Baars y Franklin, 2003)
Este modelo de MT aporta una descripción funcional básica de cómo las representaciones mentales permanecen en línea a lo largo del complejo procesamiento cognitivo (Baddeley, 2017). Tanto el bucle fonológico como la agenda visoespacial participan en el almacenamiento de la información visual y verbal. Se conciben como búferes, contenedores de información altamente procesada que no intervienen en el análisis perceptivo de la información sensorial (Buchsbaum, 2013). Ambos son controlados y supervisados a través de un mecanismo de control cognitivo conocido como ejecutivo central. La agenda visoespacial también se describe como una unidad de almacenamiento de información (Logie y Pearson, 1997). El bucle fonológico está compuesto por el almacén fonológico y el proceso de repaso articulatorio.
El bucle fonológico puede almacenar información basada en el habla durante un breve período de tiempo antes de que decaiga. La función del repaso articulatorio es evitar este decaimiento de la información, refrescando periódicamente el contenido del bucle fonológico mediante el repaso subvocal (Buchsbaum, 2013).
Por otro lado, el búfer episódico está implicado en la unión de la información, aunque las piezas aisladas multimodales de información (objeto en movimiento) se experimentan a través de diferentes canales, el búfer episódico permite la percepción de los estímulos como un todo. En algún momento, las representaciones convergen y se experimentan conscientemente como un único evento (Angelopoulou y Drigas, 2021). Por lo tanto, el mecanismo más importante para recuperar la información del búfer episódico es la percepción consciente (Baddeley, 2000). De este modo, la unión se ha convertido en la ventaja biológica más importante de la consciencia (Baddeley, 2003).
En consecuencia, la interacción entre el ejecutivo central y el búfer episódico, por un lado, y el bucle fonológico y la agenda visoespacial, por otro, está mediada por el procesamiento consciente. El ejecutivo central controla y modifica la atención, mientras que el búfer episódico permite la disponibilidad de información multimodal (Baars, 2005).
Aleksander, I. y Gamez, D. (2011). Informational Theories of Consciousness: A Review and Extension. Advances in Experimental Medicine and Biology, 718(1), 139-147- https://doi.org/10.1007/978-1-4614-0164-3_12
Alloway., T., Gathercole, S. E., Adams, A. M., Willis, C., Eaglen, R. y Lamont, E. (2005). Working Memory and Phonological Awareness as Predictors of Progress Towards Early Learning Goals at School Entry. British Journal of Developmental Psychology, 23(3), 417–426. https://doi.org/10.1348/026151005X26804
Andrade, J. (Ed). (2002). Working Memory in Perspective. Taylor Fracis Group. https://doi.org/10.4324/9780203194157
Andrs, P. y Van der Linden, M. (2001). Supervisory Attentional System in Patients with Focal Frontal Lesions. Journal of Clinical and Experimental Neuropsychology, 23(2), 225-239. https://doi.org/10.1076/jcen.23.2.225.1212
Angelopoulou, E. y Drigas, A. (2021). Working Memory, Attention and their Relationship: A theoretical Overview. Research. Society and Development,10(5), 1-8. https://doi.org/10.33448/rsd-v10i5.15288
Atkinson, R. C. y Shiffrin, R. M. (2016). Human Memory: A Proposed System and its Control Processes. En R.J. Sternberg, S.T Fiske y D.J., Foss (Eds.), Scientists Making a Difference: One Hundred Eminent Behavioral and Brain Scientists Talk about their Most Important Contributions. (pp. 115-118). Cambridge University Press. https://doi.org/10.1017/CBO9781316422250.025
Awh, E., Belopolsky, A. V. y Theeuwes, J. (2012). Top-Down versus Bottom-up Attentional Control: A Failed Theoretical Dichotomy. Trends in Cognitive Sciences, 16(8), 437-443. https://doi.org/10.1016/j.tics.2012.06.010
Baars, B. J. (1988). A Cognitive Theory of Consciousness. In A Cognitive Theory of Consciousness. Cambridge University Press.
Baars, B. J. (2005). Global Workspace Theory of Consciousness: Toward a Cognitive Neuroscience of Human Experience. Progress in Brain Research, 150(1), 45-53. https://doi.org/10.1016/S0079-6123(05)50004-9
Baars, B. J. y Franklin, S. (2003). How Conscious Experience and Working Memory Interact. Trends in Cognitive Sciences, 7(4), 166-172. https://doi.org/10.1016/S1364-6613(03)00056-1
Baddeley, A. (1992). Working Memory. Science, 255(5044), 556–559. https://doi.org/10.1126/science.1736359
Baddeley, A. (2000). The Episodic Buffer: A New Component of Working Memory? Trends in Cognitive Sciences, 4(11), 417–423. https://www.cell.com/trends/cognitive-sciences/fulltext/S1364-6613(00)01538-2?_returnURL=https%3A%2F%2Flinkinghub.elsevier.com%2Fretrieve%2Fpii%2FS1364661300015382%3Fshowall%3Dtrue
Baddeley, A. (2003). Working Memory: Looking Back and Looking Forward. Nature Reviews. Neuroscience, 4(10), 829–839. https://doi.org/10.1038/nrn1201
Baddeley, A. (2007). Working Memory, Thought, and Action. Oxford Psychology Series. https://books.google.com.co/books?hl=es&lr=&id=DRIeAAAAQBAJ&oi=fnd&pg=PT14&dq=).+Working+Memory,+Thought,+and+Action.+In+Working+memory,+thought,+and+action&ots=XsB0L2tJc7&sig=vZTNVXC-48z8Hz-kmZSOKnm88xM#v=onepage&q=).%20Working%20Memory%2C%20Thought%2C%20and%20Action.%20In%20Working%20memory%2C%20thought%2C%20and%20action&f=false
Baddeley, A. (2010). Working memory. Current Biology, 20(4). https://doi.org/10.1016/j.cub.2009.12.014
Baddeley, A. (2017). Exploring Working Memory. Taylor and Francis Group. https://doi.org/10.4324/9781315111261
Baddeley, A., Allen, R. J. y Hitch, G. J. (2010). Investigating the Episodic Buffer. Psychologica Belgica, 50(3-4), 223-243. https://doi.org/10.5334/pb-50-3-4-223
Baumeister, R. F. y Masicampo, E. J. (2010). Conscious Thought is for Facilitating Social and Cultural Interactions: How Mental Simulations Serve the Animal-culture Interface. Psychological Review, 117(3), 945-971. https://doi.org/10.1037/a0019393
Bergström, F. y Eriksson, J. (2014). Maintenance of Non-consciously Presented Information Engages the Prefrontal Cortex. Frontiers in Human Neuroscience, 8, 938. https://doi.org/10.3389/fnhum.2014.00938
Broadway, J. M. y Engle, R. W. (2011). Individual Differences in Working Memory Capacity and Temporal Discrimination. PLoS ONE, 6(10), 1-9. https://doi.org/10.1371/journal.pone.0025422
Brown, E. N., Lydic, R. y Schiff, N. D. (2010). General Anesthesia, Sleep, and Coma. New England Journal of Medicine, 363,2638-2650. https://doi.org/10.1056/NEJMra0808281
Buchsbaum, B. R. (2013). The Role of Consciousness in the Phonological Loop: Hidden in Plain Sight. Frontiers in Psychology, 4(1), 1-5. https://doi.org/10.3389/fpsyg.2013.00496
Burgess, P. W. y Stuss, D. T. (2017). Fifty Years of Prefrontal Cortex Research: Impact on Assessment. Journal of the International Neuropsychological Society, 23(9-10), 755-767. https://doi.org/10.1017/S1355617717000704
Carrasco, M. (2011). Visual Attention: The past 25 years. Vision Research, 51(13), 1484-1525. https://doi.org/10.1016/j.visres.2011.04.012
Chica, A. B., Bartolomeo, P. y Lupiáñez, J. (2013). Two Cognitive and Neural Systems for Endogenous and Exogenous Spatial Attention. Behavioural Brain Research, 237(1),107-123. https://doi.org/10.1016/j.bbr.2012.09.027
Corbetta, M. y Shulman, G. L. (2002). Control of Goal-directed and Stimulus-driven Attention in the Brain. Nature Reviews Neuroscience, 3(3), 201-215. https://doi.org/10.1038/nrn755
Corbetta, M., Patel, G. y Shulman, G. L. (2008). The Reorienting System of the Human Brain: From Environment to Theory of Mind. Neuron, 58(3), 306-324. https://doi.org/10.1016/j.neuron.2008.04.017
Cowan, N. (2012). An Embedded-Processes Model of Working Memory. En A., Miyake y P. Shah (Eds.), Models of Working Memory: Mechanisms of Active Maintenance and Executive Control (pp. 62-101). Cambridge University Press. https://doi.org/10.1017/cbo9781139174909.006
Crick, F. y Koch, C. (1990). Towards a Neurobiological Theory of Consciousness. Seminars in the Neurosciences. Seminars in the Neurosciences, 2(1), 263-265. https://authors.library.caltech.edu/40352/1/148.pdf
Crick, F. y Koch, C. (2003). A Framework for Consciousness. Nature Neuroscience, 6(2), 119-126. https://doi.org/10.1038/nn0203-119
De Loof, E., Verguts, T., Fias, W. y Van Opstal, F. (2013). Opposite Effects of Working Memory on Subjective Visibility and Priming. Journal of Experimental Psychology: Learning Memory and Cognition, 39(6), 1959-1965. https://doi.org/10.1037/a0033093
Dehaene, S. y Changeux, J. P. (2011). Experimental and Theoretical Approaches to Conscious Processing. Neuron, 70(2), 200-227. https://doi.org/10.1016/j.neuron.2011.03.018
Dehaene, S. y Naccache, L. (2001).Towards a Cognitive Neuroscience of Consciousness: Basic Evidence and a Workspace Framework. Cognition, 79(1-2), 1-37. https://doi.org/10.1016/S0010-0277(00)00123-2
Del Cul, A., Baillet, S. y Dehaene, S. (2007). Brain Dynamics Underlying the Nonlinear Threshold for Access to Consciousness. PLoS Biology, 5(10). https://doi.org/10.1371/journal.pbio.0050260
Duncan, J. (2013). The Structure of Cognition: Attentional Episodes in Mind and Brain. Neuron, 80(1), 35-50. https://doi.org/10.1016/j.neuron.2013.09.015
Earl, B. (2014). The Biological Function of Consciousness. Frontiers in Psychology, 5(1), 1-18. https://doi.org/10.3389/fpsyg.2014.00697
Eimer, M. y Grubert, A. (2014). Spatial Attention Can Be Allocated Rapidly and in Parallel to New Visual Objects. Current Biology, 24(2), 193-198. https://doi.org/10.1016/j.cub.2013.12.001
Engle, R. W. (2002). Working Memory Capacity as Executive Attention. Current Directions in Psychological Science, 11(1), 19-23. https://doi.org/10.1111/1467-8721.00160
Fingelkurts, A. A. y Fingelkurts, A. A. (2017). Information Flow in the Brain: Ordered Sequences of Metastable States. Information, 8(22), 1-9. https://doi.org/10.3390/info8010022
Fingelkurts, A. A., Fingelkurts, A. A. y Neves, C. F. H. (2010). Natural World Physical, Brain Operational, and Mind Phenomenal Space-time. Physics of Life Reviews, 7(2), 195-249. https://doi.org/10.1016/j.plrev.2010.04.001
Fingelkurts, A., Fingelkurts, A., Krause, C., Kaplan, A., Borisov, S. y Sams, M. (2003). Structural (Operational) Synchrony of EEG Alpha Activity During an Auditory Memory Task. NeuroImage, 20(1), 529-542. https://doi.org/10.1016/S1053-8119(03)00305-7
Fries, P., Roelfsema, P. R., Engel, A. K., König, P. y Singer, W. (1997). Synchronization of Oscillatory Responses in Visual Cortex Correlates with Perception in Interocular Rivalry. Proceedings of the National Academy of Sciences of the United States of America, 94(23), 12699-12704. https://doi.org/10.1073/pnas.94.23.12699
Frith, C. (2005). The Quest for Consciousness: A Neurobiological Approach. American Journal of Psychiatry, 162(2), 407. https://doi.org/10.1176/appi.ajp.162.2.407
Gayet, S., Paffen, C. L. E. y Van der Stigchel, S. (2013). Information Matching the Content of Visual Working Memory Is Prioritized for Conscious Access. Psychological Science, 24(12), 2472-2480. https://doi.org/10.1177/0956797613495882
Gray, C. M., König, P., Engel, A. K. y Singer, W. (1989). Oscillatory Responses in Cat Visual Cortex Exhibit Inter-columnar Synchronization which Reflects Global Stimulus Properties. Nature, 338(6213), 334-337. https://doi.org/10.1038/338334a0
Hassin, R. R., Bargh, J. A., Engell, A. D. y McCulloch, K. C. (2009). Implicit Working Memory. Consciousness and Cognition, 18(3), 665-678. https://doi.org/10.1016/j.concog.2009.04.003
Herculano-Houzel, S., Munk, M. H. J., Neuenschwander, S. y Singer, W. (1999). Precisely Synchronized Oscillatory Firing Patterns Require Electroencephalographic Activation. Journal of Neuroscience, 19(10), 3992-4010. https://www.jneurosci.org/content/19/10/3992
Hermes, D., Miller, K. J., Wandell, B. A. y Winawer, J. (2015). Stimulus Dependence of Gamma Oscillations in Human Visual Cortex. Cerebral Cortex, 25(9), 2951-2959. https://doi.org/10.1093/cercor/bhu091
Imas, O. A., Ropella, K. M., Ward, B. D., Wood, J. D. y Hudetz, A. G. (2005). Volatile Anesthetics Disrupt Frontal-posterior Recurrent Information Transfer at Gamma Frequencies in Rats. Neuroscience Letters, 387(3), 145-150. https://doi.org/10.1016/j.neulet.2005.06.018
Jiang, Y., Costello, P. y He, S. (2007). Processing of Invisible Stimuli: Advantage of Upright Faces and Recognizable Words in Overcoming Interocular Suppression. Psychological Science, 18(4), 349-355. https://doi.org/10.1111/j.1467-9280.2007.01902.x
Jonkisz, J. (2016). Subjectivity: A Case of Biological Individuation and an Adaptive Response to Informational Overflow. Frontiers in Psychology, 7(1), 1-6. https://doi.org/10.3389/fpsyg.2016.01206
Kertai, M. D., Whitlock, E. L. y Avidan, M. S. (2012). Brain Monitoring with Electroencephalography and the Electroencephalogram–Derived Bispectral Index During Cardiac Surgery. Anesthesia and Analgesia, 114(3),533-546. https://doi.org/10.1213/ANE.0b013e31823ee030
Koch, C., Massimini, M., Boly, M. y Tononi, G. (2016). Neural Correlates of Consciousness: Progress and Problems. Nature Reviews Neuroscience, 17(5), 307-321. https://doi.org/10.1038/nrn.2016.22
Kolb, B. y Whishaw, I.Q. (2006). Fundamentals of Human Neuropsychology. Seminars in Hearing, 27(3), 127,135. https://doi.org/10.1055/s-2006-947280
Laureys, S., Owen, A. M. y Schiff, N. D. (2004). Brain Function in Coma, Vegetative State, and Related Disorders. Lancet Neurology, 3(9), 537,546. https://doi.org/10.1016/S1474-4422(04)00852-X
Lavie, N. (2005). Distracted and Confused?: Selective Attention Under Load. Trends in Cognitive Sciences, 9(2), 75-82. https://doi.org/10.1016/j.tics.2004.12.004
Libet, B., Wright, E. W. y Gleason, C. A. (1982). Readiness-potentials Preceding Unrestricted “Spontaneous” vs. Pre-planned Voluntary Acts. Electroencephalography and Clinical Neurophysiology, 54(3), 322-335. https://doi.org/10.1016/0013-4694(82)90181-X
Lisman, J. E. y Jensen, O. (2013). The Theta-Gamma Neural Code. Neuron, 77(6), 1002-1016. https://doi.org/10.1016/j.neuron.2013.03.007
Logie, R. H. y Pearson, D. G. (1997). The Inner Eye and the Inner Scribe of Visuo-spatial Working Memory: Evidence from Developmental Fractionation. European Journal of Cognitive Psychology, 9(3), 241-257. https://doi.org/10.1080/713752559
Luo, Q., Mitchell, D., Cheng, X., Mondillo, K., McCaffrey, D., Holroyd, T., Carver, F.,Coppola, R. y Blair, J. (2009). Visual Awareness, Emotion, and Gamma band Synchronization. Cerebral Cortex, 19(8). 1896-1904. https://doi.org/10.1093/cercor/bhn216
Marchetti, G. (2014). Attention and Working Memory: Two Basic Mechanisms for Constructing Temporal Experiences. Frontiers in Psychology, 5(1), 1-15. https://doi.org/10.3389/fpsyg.2014.00880
Marchetti, G. (2018). Consciousness: A Unique Way of Processing Information. Cognitive Processing, 19(3), 435-464. https://doi.org/10.1007/s10339-018-0855-8
Melloni, L., Molina, C., Pena, M., Torres, D., Singer, W. y Rodriguez, E. (2007). Synchronization of Neural Activity across Cortical Areas Correlates with Conscious Perception. Journal of Neuroscience, 27(11), 2858-2865. https://doi.org/10.1523/JNEUROSCI.4623-06.2007
Melloni, L., Schwiedrzik, C. M., Müller, N., Rodriguez, E. y Singer, W. (2011). Expectations Change the Signatures and Timing of Electrophysiological Correlates of Perceptual Awareness. Journal of Neuroscience, 31(4), 1386-1396. https://doi.org/10.1523/JNEUROSCI.4570-10.2011
Monto, S. (2012). Nested Synchrony – A Novel Cross-scale Interaction Among Neuronal Oscillations. Frontiers in Physiology, 3(1), 1-7. https://doi.org/10.3389/fphys.2012.00384
Morsella, E., Berger, C. C. y Krieger, S. C. (2011). Cognitive and Neural Components of the Phenomenology of Agency. Neurocase, 17(3), 209-230. https://doi.org/10.1080/13554794.2010.504727
Moruzzi, G. y Magoun, H. W. (1949). Brain Stem Reticular Formation and Activation of the EEG. Electroencephalography and Clinical Neurophysiology, 1(1-4), 455-473. https://doi.org/10.1016/0013-4694(49)90219-9
Murphy, M., Bruno, M. A., Riedner, B. A., Boveroux, P., Noirhomme, Q., Landsness, E. C., Brichant, J.F., Phillips, C., Massimini, M. Laureys, S., Tononni, G. y Boly, M. (2011). Propofol Anesthesia and Sleep: A High-density EEG Study. Sleep, 34(3), 283-291. https://doi.org/10.1093/sleep/34.3.283
Oberauer, K. (2009). Design for a Working Memory. Psychology of Learning and Motivation, 51(1), 45–100. https://doi.org/10.1016/S0079-7421(09)51002-X
Pan, Y., Lin, B., Zhao, Y. y Soto, D. (2014). Working Memory Biasing of Visual Perception without Awareness. Attention, Perception, and Psychophysics, 76(7), 2051-2062. https://doi.org/10.3758/s13414-013-0566-2
Picton, T. W., Stuss, D. T., Alexander, M. P., Shallice, T., Binns, M. A. y Gillingham, S. (2007). Effects of focal frontal lesions on response inhibition. Cerebral Cortex, 17(4), 826-838. https://doi.org/10.1093/cercor/bhk031
Pitts, M. A., Metzler, S. y Hillyard, S. A. (2014). Isolating Neural Correlates of Conscious Perception from Neural Correlates of Reporting one’s Perception. Frontiers in Psychology, 5(1), 1-16. https://doi.org/10.3389/fpsyg.2014.01078
Pockett, S. y Holmes, M. D. (2009). Intracranial EEG Power Spectra and Phase Synchrony during Consciousness and Unconsciousness. Consciousness and Cognition, 18(4), 1049-1055. https://doi.org/10.1016/j.concog.2009.08.010
Purdon, P. L., Pierce, E. T., Mukamel, E. A., Prerau, M. J., Walsh, J. L., Wong, K. F. K., Salazar-Gómez, A.F., Harrell, P.G., Sampson, A.L., Cimenser, A., Ching, S., Kopell, N., Tavares-Stoeckel, C., Habeeb, K., Merhar, R. y Brown, E. N. (2013). Electroencephalogram Signatures of Loss and Recovery of Consciousness from Propofol. Proceedings of the National Academy of Sciences of the United States of America, 110(12), 1142-1151. https://doi.org/10.1073/pnas.1221180110
Railo, H., Koivisto, M. y Revonsuo, A. (2011). Tracking the Processes behind Conscious Perception: A Review of Event-related Potential Correlates of Visual Consciousness. Consciousness and Cognition, 20(3), 972-983. https://doi.org/10.1016/j.concog.2011.03.019
Ray, S. y Maunsell, J. H. R. (2011). Network Rhythms Influence the Relationship between Spike- triggered Local Field Potential and Functional Connectivity. Journal of Neuroscience, 31(35), 12674-12682. https://doi.org/10.1523/JNEUROSCI.1856-11.2011
Rodriguez, E., George, N., Lachaux, J. P., Martinerie, J., Renault, B. y Varela, F. J. (1999). Perception’s shadow: Long-distance Synchronization of Human Brain Activity. Nature, 397(6718), 430-433. https://doi.org/10.1038/17120
Roelfsema, P. R., Engel, A. K., König, P. y Singer, W. (1997). Visuomotor Integration is Associated with Zero Time-lag Synchronization Among Cortical Areas. Nature, 385(6612), 157-161. https://doi.org/10.1038/385157a0
Schiff, N. D., Nauvel, T. y Victor, J. D. (2014). Large-scale Brain Dynamics in Disorders of Consciousness. Current Opinion in Neurobiology, 25(1), 7-14. https://doi.org/10.1016/j.conb.2013.10.007
Schraw, G. y Dennison, R. S. (1994). Assessing Metacognitive Awareness. Contemporary Educational Psychology, 19(4), 460-475. https://doi.org/10.1006/ceps.1994.1033
Schraw, G. y Moshman, D. (1995). Metacognitive Theories. Educational Psychology Review, 7(4), 351-371. https://doi.org/10.1007/BF02212307
Sergent, C., Baillet, S. y Dehaene, S. (2005). Timing of the Brain Events Underlying Access to Consciousness during the Attentional Blink. Nature Neuroscience, 8(10), 1391-1400. https://doi.org/10.1038/nn1549
Shepard, R. N. y Chipman, S. (1970). Second-order Isomorphism of Internal Representations: Shapes of states. Cognitive Psychology, 1(1), 1-17. https://doi.org/10.1016/0010-0285(70)90002-2
Siclari, F., LaRocque, J. J., Postle, B. R. y Tononi, G. (2013). Assessing Sleep Consciousness within Subjects Using a Serial Awakening Paradigm. Frontiers in Psychology, 4(1), 1-9. https://doi.org/10.3389/fpsyg.2013.00542
Silverstein, B. H., Snodgrass, M., Shevrin, H. y Kushwaha, R. (2015). P3b, Consciousness, and Complex Unconscious Processing. Cortex, 73(1), 216-227. https://doi.org/10.1016/j.cortex.2015.09.004
Singer, W. (1999). Time as Coding Space? Current Opinion in Neurobiology, 9(2), 189-194. https://doi.org/10.1016/S0959-4388(99)80026-9
Smart, J. C. C. (1995). Sensations and Brain Processes. Behavioural Brain Research, 71(1-2), 157-161. https://doi.org/10.1016/0166-4328(95)00033-X
Soto, D., Mäntylä, T. y Silvanto, J. (2011). Working Memory without Consciousness. Current Biology, 21(22). https://doi.org/10.1016/j.cub.2011.09.049
Steriade, M. (2000). Corticothalamic Resonance, States of Vigilance and Mentation. Neuroscience, 101(2), 243-276. https://doi.org/10.1016/S0306-4522(00)00353-5
Steriade, M., Timofeev, I. y Grenier, F. (2001). Natural Waking and Sleep States: A View from Inside Neocortical Neurons. Journal of Neurophysiology, 85(5),1969,1985. https://doi.org/10.1152/jn.2001.85.5.1969
Stuss, D. (2011). Functions of the Frontal Lobes: Relation to Executive Functions. Journal of the International Neuropsychological Society,17 (5), 759-765. https://doi.org/10.1017/S1355617711000695
Stuss, D. T. (1991). Self, Awareness, and the Frontal Lobes: A Neuropsychological Perspective. En J. Strauss y G.R., Goethals (Eds.), The Self: Interdisciplinary Approaches (pp. 255-278). https://doi.org/10.1007/978-1-4684-8264-5_13
Stuss, D. T. y Alexander, M. P. (2005). Does Damage to the Frontal Lobes Produce Impairment in Memory? Current Directions in Psychological Science, 14(2), 84-88. https://journals.sagepub.com/doi/abs/10.1111/j.0963-7214.2005.00340.x
Stuss, D. T. y Knight, R. T. (Eds). (2009). Principles of Frontal Lobe Function. Oxford Scholarship Online. https://doi.org/10.1093/acprof:oso/9780195134971.001.0001
Sutton, S., Braren, M., Zubin, J. y John, E. R. (1965). Evoked-potential Correlates of Stimulus Uncertainty. Science, 150(3700), 1187-1188. https://doi.org/10.1126/science.150.3700.1187
Tononi, G. (2012). Integrated Information Theory of Consciousness: An Updated Account. Archives Italiennes de Biologie, 150(2-3). http://architalbiol.org/index.php/aib/article/view/15056
Tononi, G. y Koch, C. (2015). Consciousness: Here, There and Everywhere? Philosophical Transactions of the Royal Society, 370(1668), 2-18. https://doi.org/10.1098/rstb.2014.0167
Tononi, G., Boly, M., Massimini, M. y Koch, C. (2016). Integrated Information Theory: From Consciousness to its Physical Substrate. Nature Reviews Neuroscience, 17(7), 450-461. https://doi.org/10.1038/nrn.2016.44
Tsuchiya, N. y Koch, C. (2005). Continuous Flash Suppression Reduces Negative Afterimages. Nature Neuroscience, 8(8), 1096-1101. https://doi.org/10.1038/nn1500
Unsworth, N. y Engle, R. W. (2007). The Nature of Individual Differences in Working Memory Capacity: Active Maintenance in Primary Memory and Controlled Search from Secondary Memory. Psychological Review, 114, 104-132. https://doi.org/10.1037/0033-295X.114.1.104
Velichkovsky, B. B. (2017). Consciousness and Working Memory: Current Trends and Research Perspectives. Consciousness and Cognition, 55(1), 35-45. https://doi.org/10.1016/j.concog.2017.07.005
Wyart, V. y Tallon-Baudry, C. (2008). Neural Dissociation Between Visual Awareness and Spatial Attention. Journal of Neuroscience, 28(10), 2667-2679. https://doi.org/10.1523/JNEUROSCI.4748-07.2008
Zeman, A. (1996). Attentional Processing. The Brain’s Art of Mindfulness. Journal of Neurology, Neurosurgery & Psychiatry, 60(5). 596. https://doi.org/10.1136/jnnp.60.5.596
1 Artículo de reflexión derivado del proyecto de investigación: «Efecto de un entrenamiento cognitivo computarizado de memoria de trabajo sobre las actividades instrumentales de la vida diaria». Proyecto registrado en la Universidad de San Buenaventura (Medellín), 2019.
2 Magíster en Neuropsicología. Estudiante de Doctorado en Psicología Universidad San Buenaventura– Medellín. Docente programa de Psicología-Universidad Católica Luis Amigó. Carrera 22 N° 67A - 49. Manizales–Colombia. Docente programa de Medicina-Universidad de Manizales. Correo electrónico: daniel.landinezma@amigo.edu.co
3 Doctor en Psicología con orientación en Neurociencias Cognitivas Aplicadas. Docente Universidad Pontificia Bolivariana. Docente Doctorado en Psicología-Universidad San Buenaventura–Medellín. Carrera 56C # 51-110 Centro. Correo electrónico: david.montoya@usbmed.edu.co
4 Grupo de Neuropsicología y Conducta (GRUNECO). Profesor honorario de la facultad de Psicología, Universidad de San Buenaventura. Profesor Titular de Cátedra de la Facultad de Medicina, Universidad de Antioquia. Investigador emérito de Colciencias. Medellín, Colombia. Correo electrónico: david.pineda@usbmed.edu.co