
Cómo citarGómez, A. S et al ., (2024). Reconocimiento de rostros y variabilidad cultural. Análisis de la evidencia empírica y las medidas de evaluación. Ánfora, 31 (57), 299 - 325 . https://doi.org/10.30854/anf.v31.n57.2024.1102 |
Anyerson Stiths Gómez-Tabares2 https://orcid.org/0000-0001-7389-3178 CvLAC https://scienti.minciencias.gov.co/cvlac/visualizador/generarCurriculoCv.do?cod_rh=0001583946 Colombia
Jainer Amézquita Londoño3 https://orcid.org/0000-0002-7844-6753 CvLAC https://scienti.minciencias.gov.co/cvlac/visualizador/generarCurriculoCv.do?cod_rh=0001482946 Colombia
David Antonio Pineda Salazar4 https://orcid.org/0000-0002-1080-4796 CvLAC https://scienti.minciencias.gov.co/cvlac/visualizador/generarCurriculoCv.do?cod_rh=0000045640 Colombia
|
Objetivo: revisar la literatura científica que utiliza tareas cognitivas controladas y registro de actividad neurológica para evaluar la capacidad para reconocer rostros, considerando el «efecto de la otra raza» (EOR). Metodología: artículo de reflexión, cuya metodología parte de una revisión de la literatura; se incluyeron 15 estudios para la meta-síntesis. Resultados: se encontró que predominan las tareas cognitivas controladas de recuerdo subjetivo y técnica de electroencefalografía, y potenciales relacionados con eventos en la investigación sobre el reconocimiento de rostros, considerando el EOR. Se halló que la oxitocina no influye en la memoria facial, y que las dificultades en reconocer caras borrosas de otras razas correlacionan con la activación del área fusiforme facial (AFF). Conclusiones: el procesamiento neuronal de rostros de otras razas requiere más esfuerzo, evidenciado por mayor amplitud del componente N250, y relacionado con la N170. Además, invertir rostros de la propia raza prolonga su reconocimiento. La instrucción puede incrementar el procesamiento de caras de otras razas, mientras que la ira no mejora su memoria facial. Esta revisión confirma que tanto la neurofisiología como los factores culturales juegan un papel crucial en el reconocimiento facial, y sugiere que el EOR puede ser un producto de la interacción entre estos factores.
Palabras clave: reconocimiento facial; efecto de la otra raza (EOR); tareas cognitivas controladas; electroencefalografía (EEG); neurociencia cultural (obtenidos del tesauro DeCS/MeSH Descriptores en Ciencias de la Salud).
Objective: To review scientific literature in which controlled cognitive tasks and recording of neurological activity are utilized to evaluate face recognition ability, considering the "other-race effect" (ORE). Methodology: Reflection article, whose methodology is based on a literature review; 15 studies were included for meta-synthesis. Results: It was found that subjective-recollection controlled cognitive tasks, electroencephalography technique, and event-related potentials predominate in face recognition research, considering the ORE. It was also found that oxytocin has no influence on face memory, and that difficulties in recognizing other-race blurred faces correlate with the activation of the fusiform face area (FFA). Conclusions: Neural processing of other-race faces requires more effort, evidenced by larger N250 amplitude, and it is related to N170 component. Furthermore, own-race face recognition is prolonged when these are inverted. of Other-race face processing may be increased by instruction, whereas anger does not improve other-race face memory. This review confirms that both neurophysiology and cultural factors play a crucial role in face recognition and suggests that ORE may be produced by the interaction between these factors.
Keywords: face recognition; other-race effect (ORE); controlled cognitive tasks; electroencephalography (EEG); cultural neuroscience (obtained from the thesaurus DeCS/MeSH – Health Science Descriptors)
Objetivo: Rever a literatura científica que utiliza tarefas cognitivas controladas e registro da atividade neurológica para avaliar a capacidade de reconhecer rostos, considerando o "efeito de outra raça" (EOR). Metodologia: Artigo de reflexão, cuja metodologia parte de uma revisão da literatura; foram incluídos 15 estudos para a metassíntese. Resultados: Foi descoberto que predominam as tarefas cognitivas controladas de lembrança subjetiva e a técnica de eletroencefalografia, e potenciais relacionados a eventos na pesquisa sobre o reconhecimento de rostos, considerando o EOR. Verificou-se que a oxitocina não influencia a memória facial e que as dificuldades em reconhecer rostos borrados de outras raças correlacionam-se com a ativação da área fusiforme facial (AFF). Conclusões: O processamento neuronal de rostos de outras raças requer mais esforço, evidenciado por uma maior amplitude do componente N250 e relacionado ao N170. Além disso, inverter rostos da própria raça prolonga seu reconhecimento. A instrução pode aumentar o processamento de rostos de outras raças, enquanto a raiva não melhora sua memória facial. Esta revisão confirma que tanto a neurofisiologia quanto os fatores culturais desempenham um papel crucial no reconhecimento facial e sugere que o EOR pode ser um produto da interação entre esses fatores.
Palavras-chave: reconhecimento facial; efeito de outra raça (EOR); tarefas cognitivas controladas; eletroencefalografia (EEG); neurociência cultural (obtidos do tesauro DeCS/MeSH - Descritores em Ciências da Saúde)
Los rostros son estímulos visuales que transmiten información perceptual y social (Schwartz et al., 2023; Shoham et al., 2022). La información perceptiva es relevante porque permite el reconocimiento de las características de la cara; por ejemplo, el tamaño de la nariz y boca o el color de los ojos. Esto permite que las personas realicen inferencias sociales automáticas consistentes con los atributos percibidos (Abudarham & Yovel, 2016; Schwartz y Yovel, 2019a, 2019b; Shoham et al., 2022).
La capacidad para reconocer rostros es un proceso neurocognitivo complejo vinculado al procesamiento visual y la codificación social, la segmentación de rasgos y atributos faciales percibidos, y la integración de estos para construir una representación coherente y única del rostro de una persona (Blais et al., 2021; Chua et al., 2005; Schwartz et al., 2023; Tarr y Gauthier, 2000). También, implica la identificación y la representación de características faciales específicas, y su integración en un modelo mental único de cada individuo (Blais et al., 2021).
El reconocimiento de rostros y el procesamiento de expresiones faciales son procesos cognitivos diferentes, pero relacionados, y comparten algunos mecanismos y vías cerebrales (Duchaine y Yovel, 2015; Yamamoto et al., 2020). La diferencia esencial es que el reconocimiento de rostros está más asociado con el registro de las características del rostro, y el procesamiento de expresiones faciales con la interpretación de aspectos socioemocionales (Yamamoto et al., 2020).
Por un lado, el reconocimiento de rostros comienza con la percepción de características básicas de la cara, como la forma y la textura, que son procesadas por las áreas visuales primarias y secundarias del cerebro. Luego, estas características se integran en una representación facial más compleja en la corteza temporal inferior (CTI), que incluye el área fusiforme (AF) (Kanwisher et al., 1997; Kanwisher y Yovel, 2006; Pitcher y Ungerleider, 2021; Sellal, 2022). Finalmente, esta representación facial se compara con modelos mentales almacenados en la memoria para identificar a la persona (Haxby et al., 2000; Lopatina et al., 2018).
Por otro lado, el procesamiento de expresiones faciales implica la capacidad para procesar información social y afectiva a partir de la expresión del rostro (Bigelow et al., 2022; Shoham et al., 2022; Zhen et al., 2013), e involucra la actividad del surco temporal superior posterior (STSp) y el sistema límbico (Atkinson y Adolphs, 2011; Duchaine y Yovel, 2015; Haist y Anzures, 2017). En este sentido, las capacidades de reconocimiento de rostros y procesamiento de expresiones faciales son esenciales para el éxito de las interacciones sociales, su modulación y la comunicación con los demás a lo largo de la vida.
Estudios con resonancia magnética funcional (RMf) han identificado tres regiones centrales en la corteza occipitotemporal asociadas con el reconocimiento de rostros, las cuales son el AF, la región del STSp y la región del giro occipital inferior (GOI) (Gobbini y Haxby, 2007; Karimi-Rouzbahani et al., 2021; Pitcher et al., 2014; Sellal, 2022; Zhen et al., 2013). Estas tres regiones constituyen el sistema central para el reconocimiento facial. El AF y la región STSp procesan características distintivas del rostro, como la dirección de la mirada, los movimientos de labios y la expresión facial. La región del GOI se encarga de procesar aspectos invariantes que subyacen al reconocimiento de individuos (Zhen et al., 2013). También, se ha reportado que el núcleo amigdalino y la ínsula están implicados en el procesamiento de estímulos emocionales de la expresión facial (Furl et al., 2013; Gobbini y Haxby, 2007; Pitcher et al., 2014; Sellal, 2022).
Zhen et al. (2013) y Sellal (2022) sostienen que el reconocimiento de rostros es un proceso jerárquico y eficiente que involucra múltiples redes neurales especializadas en distintos aspectos del reconocimiento facial. El proceso comienza en áreas visuales primarias, y avanza hacia áreas más especializadas. Un camino principal conecta la corteza occipital, donde se encuentra el área occipital facial (AOF), con el área fusiforme facial (AFF) en el giro fusiforme, que desempeña un papel crítico en el reconocimiento de la identidad del rostro y sus aspectos invariantes. La segunda subred conecta la circunvolución frontal media izquierda (CFM) y la circunvolución frontal inferior (CFI), relacionadas con el acceso a la información semántica de los rostros; como el nombre e información biográfica de la persona (Zhen et al., 2013).
La tercera subred incluye regiones asociadas con la percepción social del rostro, como el movimiento y orientación de la mirada, las expresiones faciales y los movimientos de los labios. Esta red se extiende desde la corteza visual primaria hasta el surco temporal superior (STS), y se le conoce como la tercera vía de reconocimiento visual (Sellal, 2022; Pitcher y Ungerleider, 2021; Shoham et al., 2022). También involucra el STSp, la corteza frontal orbital (CFO) y la corteza insular (CIN), que están especialmente sintonizadas con la expresión facial. Además, sistemas funcionales adicionales como el surco intraparietal (para el manejo de la atención espacial), la corteza auditiva primaria (percepción preléxica del habla), y el sistema límbico (percepción de emociones) están conectados al sistema central de procesamiento visual para el reconocimiento de rostros (Sellal, 2022).
Varios estudios han confirmado el papel que tiene el surco temporal superior posterior (STSp) en el procesamiento de estímulos visuales vinculados al movimiento, las expresiones faciales y la mirada (Pitcher y Ungerleider, 2021; Pitcher et al., 2020; Sliwinska et al., 2020), y en procesos que respaldan la cognición social; como la atribución intencional y la teoría de la mente (Saxe y Kanwisher, 2003). Así, la evidencia respalda un modelo neurocognitivo orientado al procesamiento jerárquico y eficiente que involucra múltiples redes neurales especializadas, en el que el procesamiento visual de rostros inicia con la identificación de características básicas y avanza a niveles de procesamiento más complejos de representación mental (Haxby et al., 2000; Zhen et al., 2013; Sellal, 2022). Sin embargo, existen interrogantes acerca de la interacción entre estas áreas y cómo se ven afectadas por factores culturales e individuales.
La investigación ha mostrado que el reconocimiento de rostros no es uniforme en todas las culturas y contextos, y que la variabilidad cultural puede afectar la forma en que se procesan y recuerdan los rostros de individuos de diferentes grupos étnicos (Liu et al., 2019; Kelly et al., 2007). Esto ha llevado a neurocientíficos a explorar el efecto de la variabilidad cultural, como los estereotipos de raza, género, etnia y los sesgos endogrupales en el procesamiento de rostros (Hugenberg et al., 2007, 2010; Liu et al., 2019).
Un claro ejemplo es el «efecto de otra raza» (EOR), que describe cómo las personas tienden a ser más precisas en el reconocimiento de rostros de su propia raza que en el de otras razas (Meissner y Brigham, 2001); lo cual ha sido documentado en estudios recientes (Schwartz et al., 2023; Stelter y Schweinberger, 2023).
Por lo tanto, es crucial el estudio sobre cómo los factores culturales y las diferencias individuales generan un efecto diferencial en las áreas cerebrales involucradas en el reconocimiento de rostros, y cómo se integran en un modelo que explique la interacción entre los distintos niveles de procesamiento y las áreas funcionales (Haxby et al., 2000; Zhen et al., 2013; Sellal, 2022). Además, es necesario investigar cómo estos hallazgos pueden ayudar a entender la plasticidad del cerebro, y la influencia de factores socioculturales en el procesamiento visual. Estos aspectos tienen implicaciones importantes para el análisis de las tareas cognitivas que se han utilizado en los estudios sobre el reconocimiento de rostros y el valor de las técnicas utilizadas como potenciales relacionados con eventos (PREs), RMf y electroencefalograma EEG.
De acuerdo con lo planteado hasta el momento, es relevante realizar un análisis crítico de la evidencia sobre el efecto de la variabilidad cultural en el procesamiento de rostros y las tareas cognitivas que se emplean en la investigación científica actual. El objetivo de este trabajo es realizar una revisión sistemática de la literatura científica sobre el reconocimiento de rostro y el EOR, considerando las tareas cognitivas controladas que se emplean en la investigación neurocientífica. En esta revisión se destacarán los siguientes aspectos: la evidencia disponible sobre las diferencias en los procesos cognitivos y neurales que subyacen en el reconocimiento de rostros según la percepción social racial, las tareas cognitivas controladas, y los métodos de neuroimagen más empleados para la investigación científica.
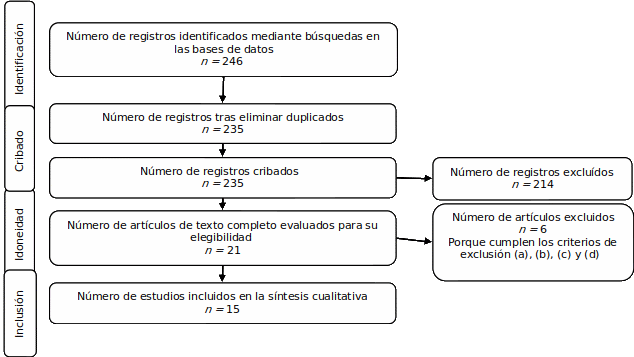
Este estudio es de reflexión, cuya metodología parte de una revisión de literatura. Se emplearon criterios de la declaración PRISMA para la elegibilidad de los estudios (Page et al., 2021). Las directrices de este sistema son ampliamente reconocidas en la comunidad científica, y se emplean para garantizar la transparencia y la calidad en los informes de las revisiones sistemáticas y los metaanálisis. A continuación, se describe el proceso de búsqueda en bases de datos, los criterios de elegibilidad y se muestra el diagrama de flujo.
Los estudios se identificaron mediante búsquedas en bases de datos electrónicas: Scopus, Springer y Science Direct. La búsqueda se realizó el 23 de abril de 2023 con la siguiente estrategia de búsqueda en inglés: "face recognition" AND "race" OR "ethnic" OR "culture" AND "eeg" OR "fmri" OR "ERPs", eligiendo únicamente artículos, para un total de 246 registros. La búsqueda no incluyó restricción temporal y se realizó por título, resumen y palabras clave.
Considerando que el objetivo es presentar la evidencia disponible en el campo de las neurociencias, y las tareas cognitivas controladas que se emplean para medir el efecto de los factores socioculturales en la capacidad para el reconocimiento de rostros, se definieron los siguientes criterios de inclusión: artículos empíricos experimentales publicados en la última década (a), cuyo tema central es el reconocimiento de rostros en humanos y el EOR (b), que utilicen medidas de funcionamiento cerebral (c) y tareas cognitivas controladas. (d) Se excluyen los artículos que: (a) poseen diseños diferentes a la investigación experimental, (b) poseen enfoque diferente a la variabilidad cultural y el EOR, (c) se desarrollan en muestras con alteraciones clínicas o animales, (d) no hacen una presentación clara del paradigma de medición de la tarea cognitiva de reconocimiento de rostros, y (e) no presentan interés en el EOR.
Cada autor asumió la búsqueda de los artículos en una de las bases de datos y, al identificar los artículos a incluir, se creó una tabla ad-hoc en la que se extrajo la información bibliométrica. Posterior a ello, se realizó una identificación conceptual de los paradigmas sobre las tareas cognitivas controladas de reconocimiento facial agrupándolos por categorías.
Con el diseño de revisión sistemática, la muestra es documental y no representa un riesgo para los humanos. Además, se ha conservado el respeto por las fuentes y los autores revisados.
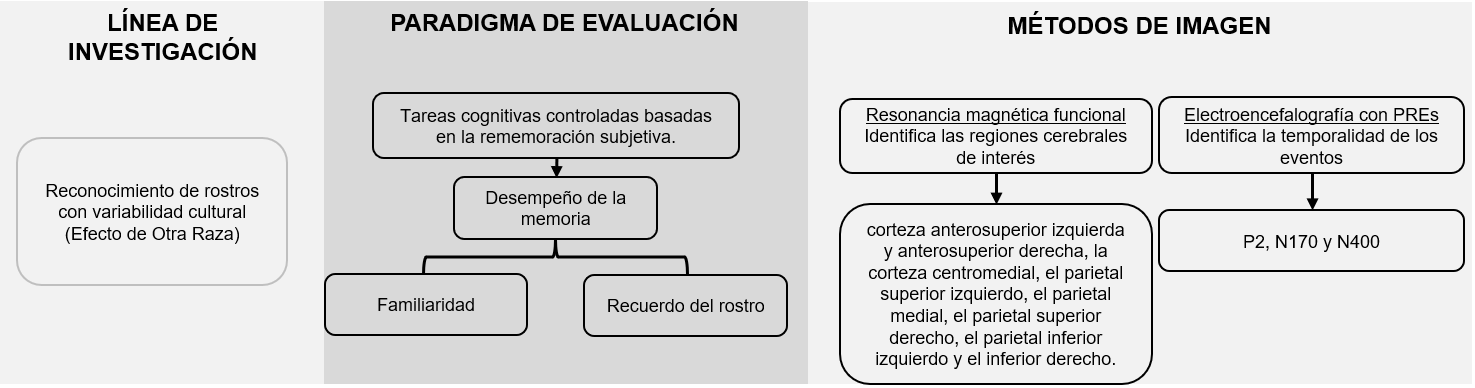
La investigación en el ámbito de la neurociencia cognitiva sobre la capacidad de reconocimiento de rostros, considerando la diversidad cultural, es una línea de estudio sólida y definida que se centra en lo que se denominó al inicio como «sesgo de otra etnia», «efecto de etnia cruzada» o «efecto de la otra raza» (EOR). Este fenómeno ha sido tradicionalmente estudiado mediante el uso de paradigmas de tareas cognitivas basadas en la rememoración subjetiva, con un enfoque específico en la evaluación del desempeño de la memoria y dos subprocesos que lo componen: la familiaridad y el recuerdo con respecto al rostro (Herzmann et al., 2013; Schwartz et al., 2023; Wong et al., 2021). Este marco metodológico facilita la comprensión de los matices, y la dinámica de los procesos cognitivos y neurales implicados en el reconocimiento de rostros en distintos contextos interculturales.
Las tareas cognitivas aplicadas en este campo de investigación, pese a presentar ciertas variaciones, generalmente implican la exposición a rostros previamente desconocidos para el sujeto experimental, seguido de una fase de aprendizaje en la que se presenta una única imagen por rostro. Posteriormente, en la fase de evaluación, estos rostros deben ser identificados entre un conjunto de imágenes distractoras nuevas (Tüttenberg y Wiese, 2019). El protocolo se completa con los participantes emitiendo juicios de rememoración-reconocimiento sobre los rostros visualizados u observados (Herzmann et al., 2017).
Existen variaciones de este paradigma diseñadas para examinar más detalladamente los procesos perceptivos y atencionales selectivos, con el objetivo de identificar las regiones cerebrales que se activan durante la detección de rostros y determinar si dicha activación es sensible a componentes faciales específicos (componentes) o a una representación holística (configuración) del rostro (Zhao et al., 2014). En estas adaptaciones del paradigma, después de cada imagen presentada, el sujeto debe emitir un juicio sobre la raza del rostro que aparece en la imagen. Estas variantes permiten un análisis más profundo de los procesos cognitivos y neurales, subyacentes al reconocimiento facial en contextos culturales diferentes.
Gran parte de la investigación neurocientífica ha integrado, de manera explícita o implícita, un modelo explicativo jerárquico para el procesamiento facial con el fin de interpretar la evidencia empírica acumulada. Este modelo se apoya en el uso de resonancia magnética funcional (RMf) para identificar el sustrato anatómico correspondiente con las diferentes etapas jerárquicas del procesamiento facial, y recurre a la electroencefalografía (EEG), particularmente los potenciales relacionados con eventos (PREs), para determinar la temporalidad precisa de estos procesos (Colombatto y McCarthy, 2017).
Los PREs representan fluctuaciones de voltaje presentes en segmentos específicos de la señal de EEG, las cuales, tras la filtración de la actividad electromuscular (por ejemplo, de los músculos maseteros y oculares), permiten visualizar campos eléctricos asociados con la actividad de conjuntos de neuronas. Estas fluctuaciones se manifiestan en forma de oscilaciones, que se denominan con base en la dirección del cambio de potencial (N1, N2 para desplazamientos negativos hacia arriba; P1, P2 para desplazamientos positivos hacia abajo) (Donchin, 1979). Estos patrones permiten un análisis riguroso y detallado de la dinámica temporal y espacial de la actividad neural durante el procesamiento facial.
La tabla 1 muestra los distintos métodos de imagen funcional empleados cuando se utilizan tareas cognitivas controladas de reconocimiento facial en diferentes poblaciones, y se detallan aspectos relacionados con su resolución, aplicación, ventajas y desventajas.
Tabla 1. Métodos de imagen funcional utilizados en tareas cognitivas controladas de reconocimiento de rostros.
|
Método de imagen |
Resolución |
Aplicación |
Ventajas |
Desventajas |
|
EEG |
Espacial- Baja Temporal- Alta |
Estudiar varios ritmos, epilepsia, mapeo preoperativo, desórdenes degenerativos. |
No invasivo, no expone a radiación iónica, ampliamente utilizada, bajo costo. |
Baja resolución espacial |
|
RMf |
Espacial- Alta Temporal- Baja |
Mapeo preoperativo y mapeo funcional |
No invasivo, puede obtener imagen funcional |
Alto costo |
Adaptado de: Brain Connectivity Analysis Methods for Better Understanding of Coupling (Shriram et al., 2012).
En la ejecución de las tareas cognitivas, los participantes interactúan con diversas tecnologías de registro de actividad cerebral. Estas están seleccionadas en función de los objetivos específicos de la investigación, para identificar la activación de las Regiones de Interés (RdI), y determinar el instante preciso de ocurrencia de procesos específicos. En el ámbito de los estudios de reconocimiento de rostros, los PREs ocupan un lugar destacado, justamente porque han contribuido a identificar tres componentes fuertemente asociados con la percepción diferencial de rostros según la pertenencia étnica o racial: la P2, la N170 y la N400 (Yong et al., 2020). Estos componentes proporcionan una visión detallada de la temporalidad y las características del procesamiento neuronal durante el reconocimiento facial, enfocándose específicamente en la respuesta a variaciones de origen étnico y racial.
Así mismo, la tabla 2 muestra la síntesis de estudios incluidos en esta revisión, en la cual se destacan las medidas neurofisiológicas, las tareas cognitivas controladas y la evidencia empírica reportada sobre la capacidad de reconocimiento de rostros, considerando el EOR. La figura 2 muestra un diagrama de síntesis analítica de los resultados presentados.
Tabla 2. Evidencia empírica sobre reconocimiento de rostros y el EOR.
|
Autores (año) |
Muestra |
Medida neurofisiológica |
Tarea cognitiva controlada |
Resultados relevantes |
|
(Herzmann et al ., 2013) |
52 adultos jóvenes, la mitad hombres (edad M 22.3, SD = 3.1) y la otra mitad mujeres (edad M 23.1, SD = 3.4). |
EEG medida con Geodesic Sensor Net de 128 canales ERPs. |
Recuerdo subjetivo Juicio recordar-saber Si/No No = familiar/no familiar |
La oxitocina no afectó la memoria de las caras de la propia raza y de otras cuando se midió entre todos los participantes, ni tuvo efecto cuando se midió para mujeres y hombres por separado. La oxitocina no afectó diferencialmente la memoria para rostros femeninos y masculinos. Mostró que los juicios de familiaridad con las caras estudiadas son más precisos después de la administración de oxitocina. |
|
(Zhao et al . , 2014) |
12 adultos jóvenes chinos, la mitad mujeres (edad M 23.7). |
fMRI Philips Achieva 3.0T System . |
Identificación de presentación repetida. |
Diferencias en el reconocimiento de la raza propia y ajena. Las caras borrosas se correlacionaron con las diferencias en la activación de AFF para esas caras, lo que sugiere que la configuración del procesamiento dentro de la AFF puede ser la base del efecto de otra raza en el reconocimiento facial. |
|
(Herzmann, 2016) |
26 adultos jóvenes caucásicos, 10 mujeres (edad M 18, SD= 0,7). |
EEG con ( Easy-CapTM ) de 32 canales.
ERPs. |
Recuerdo subjetivo Juicio recordar-saber
Grado de recuerdo de 1-4 |
Las amplitudes N250 aumentadas para rostros de otras razas se toman para representar mayores demandas neurales en el procesamiento específico de identidad de rostros de otras razas, que generalmente se procesan menos holísticamente, y menos a nivel del individuo. |
|
(Colombatto y McCarthy, 2017) |
31 adultos caucásicos, 3 hombres (edad M 23.67, SD=4,8). |
EEG con Neuroscan Quik-Cap de 64 canales. ERPs. |
Tarea de primado repetido Identificando raza B/W |
El reconocimiento de rostros de la propia raza toma más tiempo si se invierte el rostro. La inversión del rostro reveló efectos en el procesamiento, implicando áreas de la corteza visual extraestriada pericalcarina y la corteza occitotemporal lateral. |
|
(Herzmann et al., 2017) |
22 adultos jóvenes caucásicos, 73% mujeres (edad M19.1 con SD= 1.4 años). 12 orientales-asiáticos (67% mujeres (edad M 18.8 SD=1.1). |
EEG con ( easy-cap TM). ERPs. |
Tarea de memoria asociativa con recuerdo subjetivo de los momentos de codificación.
Old / New Old > Blue / Orange |
Primer estudio con este tipo de tarea para evaluar el reconocimiento de rostros. Durante la fase de estudio, los rostros de otras razas reconocidos posteriormente (con y sin información de fondo correcta) provocaron amplitudes medias más positivas que los rostros de la propia raza, lo que sugiere una mayor activación neuronal durante la codificación de rostros de otras razas. |
|
(Wiese y Schweinberger, 2018) |
40 adultos caucásicos, 20 hombres (edad M 23, SD= 2,7). |
EEG con Biosemi Active II System. ERPs. |
Recuerdo subjetivo Juicio recordar-saber Old / New |
El sesgo de la propia raza estuvo acompañado por respuestas N170 más grandes para rostros de otras razas, lo que presumiblemente refleja un procesamiento perceptivo más esforzado de esta categoría de rostros. |
|
(Herzmann et al ., 2018) |
36 adultos caucásicos, 15 hombres (edad M 21 SD=2.5). |
EEG con Geodesic Sensor NetTM de 128 canales. ERPs. |
Inversión de rostros Recuerdo subjetivo Juicio recordar-saber Grado de recuerdo de 1-4. |
Primera evidencia neuronal de que el aumento del procesamiento holístico durante la codificación de la memoria contribuye al efecto de otra raza en la memoria facial. |
|
(Tüttenberg y Wiese, 2019) |
20 adultos caucásicos, 10 mujeres (edad M 23.6 SD=5.8). |
EEG con ANT Neuro System de 64 canales. ERPs. |
3 tareas Clasificación de rostros Coincidencia Detección de objetos |
Existe una mejor clasificación y coincidencia para las identidades de la propia raza que para las de otras razas. |
|
(Proverbio et al ., 2020) |
24 adultos caucásicos, 12 hombres (edad M 23.8, SD=4,23). |
EEG y EOG 128 canales. ERPs. |
Recuerdo subjetivo Se incluyen imágenes de rostros de bebés. Juicio recordar-saber Old / New |
Un efecto de esquema de bebé en las respuestas N170, N2 anterior y P300, que eran más grandes en las caras de bebés que en las de adultos, independientemente de la etnia. |
|
(Yong et al ., 2020) |
31 adultos asiáticos, 20 mujeres (edad M 23. 18 SD= 3.33). |
EEG con Geodesic Sensor NetTM de 128 canales. ERPs. |
Identificación de raza por característica de los ojos: Caucásicos/asiáticos |
Una característica facial específica, la región de los ojos, puede explicar los efectos conocidos de la percepción de la raza en los potenciales cerebrales tempranos. |
|
(Tüttenberg y Wiese, 2021) |
36 adultos caucásicos, 26 mujeres (edad M 21.7 SD= 4.1). |
EEG con ANT Neuro System de 64 canales. ERPs. |
Recuerdo subjetivo Juicio recordar-saber Old / New |
Las instrucciones aumentan los análisis para las caras de otras razas, lo que sugiere que se asignan más recursos de procesamiento a estas caras durante la codificación. |
|
(Anzures y Mildort, 2021) |
52 adultos blancos, 29 mujeres (edad M 24.02 SD=2.04). |
EGG con Brain Vision Recorder de 64 canales. ERPs. |
Recuerdo subjetivo Juicio recordar-saber Old / New |
Las habilidades de reconocimiento de rostros y su interacción con el sesgo racial implícito modulan las primeras etapas del procesamiento de rostros de otras razas. |
|
(Kacin y Herzmann, 2021) |
47 adultos jóvenes, 27 caucásicos, (13 mujeres, edad M 19.7 SD= 1.36) 17 afroamericanos (13 mujeres, edad M 19.8, SD= 1.11). |
EEG con ( easy-cap TM ). ERPs. |
Recuerdo subjetivo Primer estudio con rostros de expresiones faciales de ira/ como coadyuvante del reconocimiento de rostros. Juicio recordar-saber Old / New |
La ira no mejoró la memoria facial de otra raza en el comportamiento de ninguna de las razas de los participantes. Evidencia de que es la experiencia con rostros de la misma raza, y no los estereotipos de rostros de otra raza, lo que influye en el EOR durante la recuperación de la memoria. |
|
(Anzures et al ., 2022) |
18 niños caucásicos, 13 mujeres (edad M 6.53 SD= 0.79). 20 niños caucásicos, 14 mujeres (edad M 9.27, SD= 0.75), y 20 adultos caucásicos, 14 mujeres (edad M 19.74 SD= 1.86). |
EEG de 64 canales con Brain Vision Recorder (Versión 1.21.0303). ERPs. |
Recuerdo subjetivo
Juicio recordar-saber
Old / New |
Las caras de otras razas provocaron amplitudes P100 más grandes que las caras de la propia raza. Además, los adultos con una mejor competencia en el reconocimiento de otras razas mostraron respuestas de amplitud P100 más grandes, en comparación con los adultos con peor competencia en el reconocimiento de otras razas. |
|
(Roth y Reynolds, 2022) |
46 bebés de 10 meses. |
EEG con
Geodesic
Sensor NetTM.
|
Primer estudio en analizar la familiarización, atención y reconocimiento facial en bebés. |
Los bebés de esta edad pueden procesar rostros humanos de manera más eficiente cuando se familiarizan con un solo ejemplar. |
El objetivo de este trabajo fue realizar una revisión crítica intencionada de la literatura científica sobre el reconocimiento de rostro y el EOR, considerando las tareas cognitivas controladas que se emplean en la investigación neurocientífica. Se hallaron diversos estudios que analizaron la relación entre el reconocimiento de rostros y la variabilidad cultural del EOR, a menudo utilizando medidas neurofisiológicas y tareas cognitivas controladas para explorar estos procesos (Anzures y Mildort, 2021; Anzures et al., 2022; Colombatto y McCarthy, 2017; Herzmann, 2016; Herzmann et al., 2013, 2017, 2018; Kacin y Herzmann, 2021; Proverbio et al., 2020; Roth y Reynolds, 2022; Tüttenberg y Wiese, 2019, 2021; Wiese y Schweinberger, 2018; Yong et al., 2020; Zhao et al., 2014).
Los estudios empíricos revisados sobre el reconocimiento de rostros, considerando el EOR, han centrado la atención en tres aspectos clave. Primero, los estímulos utilizados; típicamente imágenes de rostros caucásicos, asiáticos-orientales y afroamericanos de alta resolución, y no previamente conocidos por los participantes. Segundo, la utilización de dispositivos especializados para registrar la actividad neuronal; como RMf y EEG con PREs. Los detalles técnicos de estos dispositivos, desde las marcas y el número de canales hasta las estrategias de análisis de datos y la ubicación de los electrodos, son cuidadosamente descritos y controlados en estos estudios. El tercer elemento se refiere al procedimiento de desarrollo de la tarea cognitiva, incluyendo la presentación de las imágenes en bloques o secuencias, la recopilación de las respuestas de los participantes (generalmente a través de la presión de teclas), y las instrucciones proporcionadas. Además, estos estudios a menudo incluyen instrumentos de medición adicionales, como auto reportes.
Las tareas cognitivas controladas, utilizadas en la evaluación del reconocimiento facial, destacan por las variaciones en la manipulación de las imágenes de rostros que utilizan como estímulos; por ejemplo, en su orientación utilizando rostros invertidos (Colombatto y McCarthy, 2017) e imágenes del mismo rostro desde diferentes ángulos (Tüttenberg y Wiese, 2019). Así mismo, en su composición, agregando fondos de color en la fase de codificación (Herzmann et al., 2017); en su nitidez, presentando las imágenes con efecto borroso (Zhao et al., 2014); en su estructura, mostrando solo algunos elementos del rostro (Yong et al., 2020); o agregando expresiones faciales mediante rostros con emociones como la ira (Kacin y Herzmann, 2021).
Como características distintivas presentadas en el apartado de la muestra, se observa en los estudios la importancia de reportar el daño cerebral en la selección; así como la dominancia manual de los participantes, indicando las precisiones en la ejecución diferenciada en el uso de los teclados de los ordenadores en las tareas cognitivas (Colombatto y McCarthy, 2017; Herzmann, 2016; Herzmann et al., 2017; Kacin y Herzmann, 2021), por los efectos de la motricidad en el registro de la medición neural. Esta atención al detalle refuerza la fiabilidad y validez de las conclusiones derivadas de estos estudios, reflejando la complejidad de los procesos neuronales y culturales implicados en el reconocimiento de rostros.
La investigación neurofisiológica ha mostrado una interrelación compleja de factores que influyen en el reconocimiento de rostros al considerar aspectos como el EOR. Al respecto, Herzmann et al. (2013) encontraron que la oxitocina no afectó diferencialmente la memoria para rostros de diferentes razas ni géneros. Zhao et al. (2014) identificaron diferencias en la activación cerebral relacionadas con el reconocimiento de rostros de la propia y otra raza. Estos estudios sientan las bases para el trabajo de Herzmann (2016), Colombatto y McCarthy (2017), quienes mostraron que el procesamiento específico de la identidad y la orientación del rostro son cruciales en la percepción de rostros de diferentes razas. Por su parte, Wiese y Schweinberger (2018) profundizaron en estos hallazgos, demostrando que las respuestas neurales pueden variar en función de la raza del rostro percibido.
Los estudios con EEG y PREs señalaron que las variaciones en la respuesta neural derivadas de la percepción de rostros de otras razas se concentran especialmente en la corteza anterosuperior izquierda y anterosuperior derecha, la corteza centromedial, el parietal superior izquierdo, el parietal medial, el parietal superior derecho, el parietal inferior izquierdo y el inferior derecho. El registro de la actividad neural en estas zonas ha revelado que las frecuencias que están más asociadas con la ventaja en el reconocimiento de rostros de la propia raza, en comparación con los de otras razas, se encuentran en los potenciales N170, P2 y N400 (Wiese y Schweinberger, 2018; Yong et al., 2020). A partir de estos se pueden obtener mapas de voltaje de ondas de diferencia en los potenciales evocados por los estímulos visuales (PREs, por sus siglas en inglés) entre juicios de memoria subsiguientes. Estos mapas muestran una ventana entre 500–900 ms. para el recuerdo de rostros de la propia raza y de otras (Herzmann et al., 2018).
La N170 es un PREs registrado durante la codificación de rostros entre 150 y 190 ms. N170 es una respuesta específica del rostro que refleja el procesamiento de esquemas faciales (Proverbio et al., 2020). P2, por su parte, suele estar relacionado con procesos atencionales tempranos que a menudo se consideran automáticos, y que pueden reflejar la actividad de un sistema de vigilancia temprano dedicado a la detección de estímulos amenazantes (Yong et al., 2020). Respuestas ampliadas de P2 reflejan un procesamiento más esforzado, que provocaría una mayor retroalimentación de las áreas visuales superiores a las inferiores (Anzures y Mildort, 2021). Y N400, registrado en la ventana temporal de 300–500 ms., mide los procesos de familiaridad de los rostros; mientras que el registro posterior de 500–800 ms. refleja los procesos de recuerdo (Herzmann et al., 2018; Rugg y Curran, 2007).
El estudio de Tüttenberg y Wiese (2019) evidenció que las personas tienden a asignar más recursos de procesamiento a los rostros de su propia raza, lo que resulta en una mejor clasificación y reconocimiento facial. Estos hallazgos sugieren que existe una ventaja de aprendizaje para las identidades de la propia raza, y subraya la importancia de la experiencia perceptual en el sesgo de esta. Además, concuerdan con lo reportado por Herzmann et al. (2018), quienes señalaron que el aumento en la manipulación del procesamiento holístico influye en la codificación de la memoria para rostros de la propia raza en comparación con los de otras razas. Los estudios en poblaciones más jóvenes, como el de Anzures et al. (2022), Roth y Reynolds (2022), han añadido datos similares y complementarios al mostrar que los niños y los bebés muestran variaciones en la percepción de rostros de diferentes razas.
A partir de la atención meticulosa a los estímulos, la metodología y las consideraciones demográficas en los estudios revisados, se evidencia que los factores neurofisiológicos y socioculturales interactúan para influir en el reconocimiento de rostros (Anzures y Mildort, 2021; Anzures et al., 2022; Colombatto y McCarthy, 2017; Herzmann, 2016; Herzmann et al., 2013, 2017, 2018; Kacin y Herzmann, 2021; Proverbio et al., 2020; Roth y Reynolds, 2022; Tüttenberg y Wiese, 2019, 2021; Wiese y Schweinberger, 2018; Yong et al., 2020; Zhao et al., 2014). Estos estudios resaltan la relevancia del EOR en la investigación neurocientífica, y respaldan la idea de que las diferencias en la percepción racial pueden moldear la capacidad para el reconocimiento de rostros (Anzures et al., 2013; Ge et al., 2009; Kelly et al., 2007; Meissner y Brigham, 2001; Sangrigoli & de Schonen, 2004), y proporcionan una perspectiva de estudio interdisciplinar para analizar los mecanismos de conectividad funcional para el procesamiento de rostros desde una perspectiva transcultural (Wong et al., 2020).
Asimismo, se ha demostrado que el EOR se desarrolla tempranamente durante la infancia (Anzures et al., 2022; Kelly et al., 2007; Roth y Reynolds, 2022; Sangrigoli y de Schonen, 2004), y se asocia con la exposición limitada a rostros de otras razas durante períodos críticos del desarrollo. Esto se debe a que, desde el nacimiento, los niños tienen una mayor exposición a personas pertenecientes a su endogrupo —étnico o racial―. En consecuencia, desarrollan una mayor capacidad para identificar y procesar las características y expresiones de los rostros de su propia raza, además de exhibir una preferencia por estos en comparación con los de otras razas (Bar-Haim et al., 2006; Hugenberg et al., 2007, 2010; Kelly et al., 2007).
De acuerdo con el modelo de categorización-individuación del EOR (Hugenberg et al., 2007), los rostros pertenecientes al endogrupo y al exogrupo son objeto de atención de maneras distintas en términos cualitativos. Al respecto, Prunty et al. (2023) sostienen que las características asociadas con la identidad son procesadas con mayor facilidad en los rostros del endogrupo. Esto resulta en un mejor desempeño en tareas de memoria de reconocimiento, mientras que las características vinculadas a la categoría son atendidas de manera preferente en los rostros del exogrupo; lo cual conduce a una mayor rapidez en la categorización durante las tareas de búsqueda.
A medida que los niños crecen, esta preferencia se vuelve más pronunciada, lo que sugiere actitudes sociocognitivas de sesgo racial implícito (Lebrecht et al., 2009). La exposición temprana y continua a rostros de la propia raza promueve el EOR durante el desarrollo (Anzures et al., 2013, 2022; Kelly et al., 2007). Factores como el contacto intergrupal, la exposición social, la formación de estereotipos sociales y sesgos endogrupales pueden fortalecer los sesgos raciales implícitos en el reconocimiento de rostros de otras razas (Anzures et al., 2022; Ge et al., 2009; Prunty et al., 2023). Al respecto, Lebrecht et al. (2009) mostraron que el entrenamiento perceptivo de otras razas (personas caucásicas expuestas a rostros afroamericanos) reducía el sesgo racial implícito, demostrando una relación causal con el EOR. Este efecto también se ha demostrado en niños preescolares (Qian et al., 2019).
Lo expuesto evidencia la influencia de la exposición a entornos socioculturales específicos en el procesamiento neurocognitivo de rostros. Estos hallazgos son coherentes con lo reportado en varios de los estudios de esta revisión. Zhao et al. (2014) encontraron una correlación entre las diferencias en la activación de la AFF y el reconocimiento de caras borrosas de diferentes razas, sugiriendo una base neural del EOR. Herzmann (2016) observó que la presencia de amplitudes N250, aumentadas para rostros de otras razas, podría indicar mayores demandas neurales para el procesamiento específico de la identidad. Idea respaldada por Wiese y Schweinberger (2018) y Tüttenberg y Wiese (2021), quienes reportaron respuestas N170 más grandes para rostros de otras razas, y un incremento en análisis para estas debido a instrucciones específicas.
Por su parte, Colombatto y McCarthy (2017) y Herzmann et al. (2018) señalaron que el reconocimiento de rostros de la propia raza es más lento cuando están invertidos, implicando que un mayor procesamiento holístico durante la codificación de la memoria puede contribuir al EOR. Tüttenberg y Wiese (2019), y Anzures y Mildort (2021) expandieron este hallazgo al considerar que las capacidades altas para reconocer rostros e identidad de la propia raza en comparación con otras se relaciona con los sesgos raciales implícitos. Finalmente, Anzures et al. (2022) proporcionaron evidencia que indica que la percepción de rostros de otras razas genera amplitudes P100 más grandes que la de la propia; aspectos asociados con los procesos de categorización perceptiva y la percepción social.
Estudios adicionales muestran que las personas tienen una mejor memoria de reconocimiento de rostros de su propio grupo, en comparación con los de otros grupos étnicos o raciales (Liu et al., 2019; Zhou et al., 2021). Se cree que la categorización visual de rostros según la pertenencia a un grupo (en este caso por raza) es un componente clave que explica los cambios en el procesamiento cognitivo de rostros de acuerdo con la pertenencia cultural (Hugenberg et al., 2010; Prunty et al., 2023). Estos hallazgos tienen implicaciones importantes para la comprensión de la neurociencia del procesamiento de rostros en humanos.
Los datos muestran que el procesamiento visual de rostros puede variar según la exposición sociocultural particular, lo cual está asociado al EOR. Sin embargo, también señalan que existen mecanismos neurales universales subyacentes al reconocimiento de rostros (Blais et al., 2021; Caldara et al., 2010; Sellal, 2022; Zhen et al., 2013). Esto es coherente con estudios previos que han identificado áreas cerebrales específicas, como el área fusiforme de la cara (AFF) y el área occipitotemporal (OTS), que están involucradas en el procesamiento de rostros en personas de otras razas (Kanwisher et al., 1997; Karimi-Rouzbahani et al., 2021; Gauthier et al., 1999; Pitcher y Ungerleider, 2021; Sellal, 2022; Walker et al., 2008; Zhao et al., 2014).
Finalmente, el EOR se relaciona con las diferencias culturales en la distribución y el enfoque de la información visual. Contribuye a la comprensión de cómo se procesan y se perciben los rostros de personas de otras razas. Estas diferencias pueden tener implicaciones para la forma en que los mecanismos neurales subyacentes al reconocimiento de rostros se adaptan y se ajustan en función de la experiencia y el entorno cultural.
Los estudios sobre el reconocimiento de rostros y el EOR han utilizado variaciones de las tareas cognitivas, controladas para la medición de la memoria de rostros con registro neurofisiológico preferente de EEG con PREs, que muestran mayor interés por el hallazgo de las fases del proceso tras tener una fundamentación teórica sólida de las RdI en AFF y OTS gracias a la RMf. Dichas variaciones suelen enfocarse en la presentación de los estímulos. Además, el registro de la actividad neurofisiológica se enfoca de manera predominante en los potenciales P100, N170, P2 y N400. En general, los estudios señalan un mejor reconocimiento de rostros en el propio grupo en comparación con otros grupos raciales.
Abudarham, N., & Yovel, G. (2016). Reverse Engineering the Face Space: Discovering the Critical Features for Face Identification. Journal of Vision, 16 (3), 1-18. https://doi.org/10.1167/16.3.40
Anzures, G., & Mildort, M. (2021). Do Perceptual Expertise and Implicit Racial Bias Predict Early Face-Sensitive ERP Responses? Brain and Cognition, 147 , 1-12. https://doi.org/10.1016/j.bandc.2020.105671
Anzures, G., Mildort, M., Fennell, E., Bell, C., & Soethe, E. (2022). Race and Early Face-Sensitive Event-Related Potentials in Children and Adults. Journal of Experimental Child Psychology, 214 , 1-21. https://doi.org/10.1016/j.jecp.2021.105287
Anzures, G., Quinn, P. C., Pascalis, O., Slater, A. M., & Lee, K. (2013). Development of Own-Race Biases. Visual Cognition, 21 (9–10), 1165–1182. https://doi.org/10.1080/13506285.2013.821428
Atkinson, A. P., & Adolphs, R. (2011). The Neuropsychology of Face Perception: Beyond Simple Dissociations and Functional Selectivity. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 366 (1571), 1726–1738. https://doi.org/10.1098/rstb.2010.0349
Bar-Haim, Y., Ziv, T., Lamy, D., & Hodes, R. M. (2006). Nature and Nurture in Own-Race Face Processing. Psychological Science, 17 (2), 159–163. https://doi.org/10.1111/j.1467-9280.2006.01679.x
Bigelow, F. J., Clark, G. M., Lum, J. A. G., & Enticott, P. G. (2022). Facial Emotion Processing and Language During Early-to-Middle Childhood Development: An Event Related Potential Study. Developmental Cognitive Neuroscience, 53 , 1-12. https://doi.org/10.1016/j.dcn.2021.101052
Blais, C., Linnell, K. J., Caparos, S., & Estéphan, A. (2021). Cultural Differences in Face Recognition and Potential Underlying Mechanisms. Frontiers in Psychology, 12 , 1-8. https://doi.org/10.3389/fpsyg.2021.627026
Caldara, R., Zhou, X., & Miellet, S. (2010). Putting Culture Under the ‘Spotlight’ Reveals Universal Information Use for Face Recognition. PloS One, 5 (3), 1-12. https://doi.org/10.1371/journal.pone.0009708
Chua, H. F., Boland, J. E., & Nisbett, R. E. (2005). Cultural Variation in Eye Movements During Scene Perception. Proceedings of the National Academy of Sciences of the United States of America, 102 (35), 12629–12633. https://doi.org/10.1073/pnas.0506162102
Colombatto, C., & McCarthy, G. (2017). The Effects of Face Inversion and Face Race on the P100 ERP. Journal of Cognitive Neuroscience, 29 (4), 664–676. https://doi.org/10.1162/jocn_a_01079
Donchin, E. (1979). Event-Related Brain Potentials: A Tool in the Study of Human Information Processing. In H. Begleiter (Ed.) Evoked Brain Potentials and Behavior (pp. 13-88). Springer.
Duchaine, B., & Yovel, G. (2015). A Revised Neural Framework for Face Processing. Annual Review of Vision Science, 1 (1), 393–416. https://doi.org/10.1146/annurev-vision-082114-035518
Furl, N., Henson, R. N., Friston, K. J., & Calder, A. J. (2013). Top-Down Control of Visual Responses to Fear by the Amygdala. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 33 (44), 17435–17443. https://doi.org/10.1523/jneurosci.2992-13.2013
Gauthier, I., Tarr, M. J., Anderson, A. W., Skudlarski, P., & Gore, J. C. (1999). Activation of the Middle Fusiform “Face Area” Increases with Expertise in Recognizing Novel Objects. Nature Neuroscience, 2 (6), 568–573. https://doi.org/10.1038/9224
Ge, L., Zhang, H., Wang, Z., Quinn, P. C., Pascalis, O., Kelly, D., Slater, A., Tian, J., & Lee, K. (2009). Two Faces of the Other-Race Effect: Recognition and Categorisation of Caucasian and Chinese faces. Perception, 38 (8), 1199–1210. https://doi.org/10.1068/p6136
Gobbini, M. I., & Haxby, J. V. (2007). Neural Systems for Recognition of Familiar Faces. Neuropsychologia, 45 (1), 32–41. https://doi.org/10.1016/j.neuropsychologia.2006.04.015
Haist, F., & Anzures, G. (2017). Functional Development of the Brain’s Face ‐ Processing System. Wiley Interdisciplinary Reviews. Cognitive Science, 8 (1–2), 1-14. https://doi.org/10.1002/wcs.1423
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The Distributed Human Neural System for Face Perception. Trends in Cognitive Sciences, 4 (6), 223–233. https://doi.org/10.1016/s1364-6613(00)01482-0
Herzmann, G. (2016). Increased N250 Amplitudes for Other-Race Faces Reflect More Effortful Processing at the Individual Level. International Journal of Psychophysiology, 105 , 57–65. https://doi.org/10.1016/j.ijpsycho.2016.05.001
Herzmann, G., Bird, C. W., Freeman, M., & Curran, T. (2013). Effects of Oxytocin on Behavioral and ERP Measures of Recognition Memory for Own-Race and Other-Race Faces in Women and Men. Psychoneuroendocrinology, 38 (10), 2140–2151. https://doi.org/10.1016/j.psyneuen.2013.04.002
Herzmann, G., Minor, G., & Adkins, M. (2017). Neural Correlates of Memory Encoding and Recognition for Own-Race and Other-Race Faces in an Associative-Memory Task. Brain Research, 1655 , 194–203. https://doi.org/10.1016/j.brainres.2016.10.028
Herzmann, G., Minor, G., & Curran, T. (2018). Neural Evidence for the Contribution of Holistic Processing but not Attention Allocation to the Other-Race Effect on Face Memory. Cognitive, Affective & Behavioral Neuroscience, 18 (5), 1015–1033. https://doi.org/10.3758/s13415-018-0619-z
Hugenberg, K., Miller, J., & Claypool, H. M. (2007). Categorization and Individuation in the Cross-Race Recognition Deficit: Toward a Solution to An Insidious Problem. Journal of Experimental Social Psychology, 43 (2), 334–340. https://doi.org/10.1016/j.jesp.2006.02.010
Hugenberg, K., Young, S. G., Bernstein, M. J., & Sacco, D. F. (2010). The Categorization-Individuation Model: An Integrative Account of the Other-Race Recognition Deficit. Psychological Review, 117 (4), 1168–1187. https://doi.org/10.1037/a0020463
Kacin, M., & Herzmann, G. (2021). Facial Expressions of Anger Improve Neural Correlates of Memory Retrieval but not Encoding of Only Same-Race Faces. Neuropsychologia, 159 , 1-13. https://doi.org/10.1016/j.neuropsychologia.2021.107915
Kanwisher, N., McDermott, J., & Chun, M. M. (1997). The Fusiform Face Area: A Module in Human Extrastriate Cortex Specialized for Face Perception. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 17 (11), 4302–4311. https://doi.org/10.1523/jneurosci.17-11-04302.1997
Kanwisher, N., & Yovel, G. (2006). The Fusiform Face Area: A Cortical Region Specialized for the Perception of Faces. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 361 (1476), 2109–2128. https://doi.org/10.1098/rstb.2006.1934
Karimi-Rouzbahani, H., Ramezani, F., Woolgar, A., Rich, A., & Ghodrati, M. (2021). Perceptual Difficulty Modulates the Direction of Information Flow in Familiar Face Recognition. NeuroImage, 233 , 1-15. https://doi.org/10.1016/j.neuroimage.2021.117896
Kelly, D. J., Quinn, P. C., Slater, A. M., Lee, K., Ge, L., & Pascalis, O. (2007). The Other-Race Effect Develops During Infancy: Evidence of Perceptual Narrowing. Psychological Science, 18 (12), 1084–1089. https://doi.org/10.1111/j.1467-9280.2007.02029.x
Lebrecht, S., Pierce, L. J., Tarr, M. J., & Tanaka, J. W. (2009). Perceptual Other-Race Training Reduces Implicit Racial Bias. PloS One, 4 (1), 1-7. https://doi.org/10.1371/journal.pone.0004215
Liu, X., Liang, X., Feng, C., & Zhou, G. (2019). Self-Construal Priming Affects Holistic Face Processing and Race Categorization, but not Face Recognition. Frontiers in Psychology, 10 , 1-52. https://doi.org/10.3389/fpsyg.2019.01973
Lopatina, O. L., Komleva, Y. K., Gorina, Y. V., Higashida, H., & Salmina, A. B. (2018). Neurobiological Aspects of Face Recognition: The Role of Oxytocin. Frontiers in Behavioral Neuroscience , 12(195), 1-12. https://doi.org/10.3389/fnbeh.2018.00195
Meissner, C. A., & Brigham, J. C. (2001). Thirty Years of Investigating the Own-Race Bias in Memory for Faces: A Meta-Analytic Review. Psychology, Public Policy, and Law, 7 (1), 3–35. https://doi.org/10.1037/1076-8971.7.1.3
Page, M. J., McKenzie, J. E., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., Shamseer, L., Tetzlaff, J. M., Akl, E. A., Brennan, S. E., Chou, R., Glanville, J., Grimshaw, J. M., Hróbjartsson, A., Lalu, M. M., Li, T., Loder, E. W., Mayo-Wilson, E., McDonald, S., et al . (2021). The PRISMA 2020 Statement: an updated guideline for reporting systematic reviews. BMJ, 372 (71), 1-9. https://doi.org/10.1136/bmj.n71
Pitcher, D., Duchaine, B., & Walsh, V. (2014). Combined TMS and fMRI Reveal Dissociable Cortical Pathways for Dynamic and Static Face Perception. Current Biology: CB, 24 (17), 2066–2070. https://doi.org/10.1016/j.cub.2014.07.060
Pitcher, D., Pilkington, A., Rauth, L., Baker, C., Kravitz, D. J., & Ungerleider, L. G. (2020). The Human Posterior Superior Temporal Sulcus Samples Visual Space Differently from Other Face-Selective Regions. Cerebral Cortex, 30 (2), 778–785. https://doi.org/10.1093/cercor/bhz125
Pitcher, D., & Ungerleider, L. G. (2021). Evidence for a Third Visual Pathway Specialized for Social Perception. Trends in Cognitive Sciences, 25 (2), 100–110. https://doi.org/10.1016/j.tics.2020.11.006
Proverbio, A. M., Parietti, N., & De Benedetto, F. (2020). No Other Race Effect (ORE) for Infant Face Recognition: A Memory Task. Neuropsychologia, 141 , 1-11. https://doi.org/10.1016/j.neuropsychologia.2020.107439
Prunty, J. E., Jenkins, R., Qarooni, R., & Bindemann, M. (2023). Ingroup and Outgroup Differences in Face Detection. British Journal of Psychology, 114 (S1), 94–111. https://doi.org/10.1111/bjop.12588
Qian, M. K., Quinn, P. C., Heyman, G. D., Pascalis, O., Fu, G., & Lee, K. (2019). A Long ‐ Term Effect of Perceptual Individuation Training on Reducing Implicit Racial Bias in Preschool Children. Child Development, 90 (3), 1-16. https://doi.org/10.1111/cdev.12971
Roth, K. C., & Reynolds, G. D. (2022). Neural Correlates of Subordinate-Level Categorization of Own- and Other-Race Faces in Infancy. Acta Psychologica, 230 , 1-12. https://doi.org/10.1016/j.actpsy.2022.103733
Rugg, M. D., & Curran, T. (2007). Event-Related Potentials and Recognition Memory. Trends in Cognitive Sciences, 11 (6), 251–257. https://doi.org/10.1016/j.tics.2007.04.004
Sangrigoli, S., & de Schonen, S. (2004). Recognition of Own-Race and Other-Race Faces by Three-Month-Old Infants. Journal of Child Psychology and Psychiatry, and Allied Disciplines, 45 (7), 1219–1227. https://doi.org/10.1111/j.1469-7610.2004.00319.x
Saxe, R., & Kanwisher, N. (2003). People Thinking About Thinking People. The Role of the Temporo-Parietal Junction in “Theory of Mind”. NeuroImage, 19 (4), 1835–1842. https://doi.org/10.1016/s1053-8119(03)00230-1
Schwartz, L., Cohen, M., Xu, S., Liu, J., & Yovel, G. (2023). The Social ‐ Encoding Benefit in Face Recognition is Generalized to Other ‐ Race Faces. British Journal of Psychology, 114 (S1), 213–229. https://doi.org/10.1111/bjop.12592
Schwartz, L., & Yovel, G. (2019a). Independent Contribution of Perceptual Experience and Social Cognition to Face Recognition. Cognition, 183 , 131–138. https://doi.org/10.1016/j.cognition.2018.11.003
Schwartz, L., & Yovel, G. (2019b). Learning Faces as Concepts Rather Than Percepts Improves Face Recognition. Journal of Experimental Psychology. Learning, Memory, and Cognition, 45 (10), 1733–1747. https://doi.org/10.1037/xlm0000673
Sellal, F. (2022). Anatomical and Neurophysiological Basis of Face Recognition. Revue Neurologique, 178 (7), 649–653. https://doi.org/10.1016/j.neurol.2021.11.002
Shriram, R., Sundhararajan, D. M., & Daimiwal, N. (2012). Brain Connectivity Analysis Methods for Better Understanding of Coupling. International Journal of Computer Science and Information Security, 10 (11), 1-7. https://arxiv.org/ftp/arxiv/papers/1212/1212.3786.pdf
Shoham, A., Kliger, L., & Yovel, G. (2022). Learning Faces as Concepts Improves Face Recognition by Engaging the Social Brain Network. Social Cognitive and Affective Neuroscience, 17 (3), 290–299. https://doi.org/10.1093/scan/nsab096
Sliwinska, M. W., Elson, R., & Pitcher, D. (2020). Dual-Site TMS Demonstrates Causal Functional Connectivity Between the Left and Right Posterior Temporal Sulci During Facial Expression Recognition. Brain Stimulation, 13 (4), 1008–1013. https://doi.org/10.1016/j.brs.2020.04.011
Stelter, M., & Schweinberger, S. R. (2023). Understanding the Mechanisms Underlying the Other ‐ ‘Race’ Effect: An Attempt at Integrating Different Perspectives. British Journal of Psychology, 114 (S1), 1–9. https://doi.org/10.1111/bjop.12615
Tarr, M. J., & Gauthier, I. (2000). FFA: A flexible Fusiform Area for Subordinate-Level Visual Processing Automatized by Expertise. Nature Neuroscience, 3 (8), 764–769. https://doi.org/10.1038/77666
Tüttenberg, S. C., & Wiese, H. (2019). Learning Own- and Other-Race Facial Identities: Testing Implicit Recognition with Event-Related Brain Potentials. Neuropsychologia, 134 , 1-43. https://doi.org/10.1016/j.neuropsychologia.2019.107218
Tüttenberg, S. C., & Wiese, H. (2021). Recognising Other-Race Faces is More Effortful: The Effect of Individuation Instructions on Encoding-Related ERP Dm Effects. Biological Psychology, 158 , 1-8. https://doi.org/10.1016/j.biopsycho.2020.107992
Walker, P. M., Silvert, L., Hewstone, M., & Nobre, A. C. (2008). Social Contact and Other-Race Face Processing in the Human Brain. Social Cognitive and Affective Neuroscience, 3 (1), 16–25. https://doi.org/10.1093/scan/nsm035
Wiese, H., & Schweinberger, S. R. (2018). Inequality Between Biases in Face Memory: Event-Related Potentials Reveal Dissociable Neural Correlates of Own-Race and Own-Gender Biases. Cortex; A Journal Devoted to the Study of the Nervous System and Behavior, 101 , 119–135. https://doi.org/10.1016/j.cortex.2018.01.016
Wong, H. K., Estudillo, A. J., Stephen, I. D., & Keeble, D. R. T. (2021). The Other-Race Effect and Holistic Processing Across Racial Groups. Scientific Reports, 11 (1), 1-15. https://doi.org/10.1038/s41598-021-87933-1
Wong, H. K., Stephen, I. D., & Keeble, D. R. T. (2020). The Own-Race Bias for Face Recognition in a Multiracial Society. Frontiers in Psychology, 11 (208), 1-8. https://doi.org/10.3389/fpsyg.2020.00208
Yamamoto, H., Kawahara, M., Kret, M., & Tanaka, A. (2020). Cultural Differences in Emoticon Perception: Japanese See the Eyes and Dutch the Mouth of Emoticons. Letters on Evolutionary Behavioral Science, 11 (2), 40–45. https://doi.org/10.5178/lebs.2020.80
Yong, M. H., Lim, X. L., & Schaefer, A. (2020). How do Asians Perceive Caucasian Eyes? Electrophysiological Correlates of Perceiving Racial Differences from the Eyes Region of the Face. Neuroscience Letters, 720 , 1-6. https://doi.org/10.1016/j.neulet.2020.134759
Zhao, M., Cheung, S.-H., Wong, A. C.-N., Rhodes, G., Chan, E. K. S., Chan, W. W. L., & Hayward, W. G. (2014). Processing of Configural and Componential Information in Face-Selective Cortical Areas. Cognitive Neuroscience, 5 (3–4), 160–167. https://doi.org/10.1080/17588928.2014.912207
Zhen, Z., Fang, H., & Liu, J. (2013). The Hierarchical Brain Network for Face Recognition. PloS One, 8 (3), 1-9. https://doi.org/10.1371/journal.pone.0059886
Zhou, X., Burton, A. M., & Jenkins, R. (2021). Two Factors in Face Recognition: Whether You Know the Person’s Face and Whether You Share the Person’s Race. Perception, 50 (6), 524–539. https://doi.org/10.1177/03010066211014016
1 Producto vinculado a la Universidad Católica Luis Amigó y a la Universidad de San Buenaventura . Grupo de Investigación «Neurociencias Básicas y Aplicadas». Financiación: no recibió financiación. Declaración de intereses: los autores declaran que no existe conflicto de intereses. Disponibilidad de datos: todos los datos relevantes se encuentran en el artículo.
2 Doctor en Filosofía. Candidato a Doctor en Psicología. Docente de la Universidad Católica Luis Amigó. Correo electrónico: anyerspn.gomezta@amigo.edu.co
3 Magíster en Psicología. Candidato a Doctor en Psicología. Docente de la Universidad de San Buenaventura. Correo electrónico: jainer.amezquita@tau.usbmed.edu.co
4 Doctor en Psicología. Profesor titular de la Facultad de Psicología. Universidad de San Buenaventura. Correo electrónico: david.pineda@usbmed.edu.co